Evolution © Eric R. Pianka All life on Earth arose from one common ancestor whose descendents underwent divergent selection. Evidence for this is overwhelming: every Earthly living entity uses the same genetic code. All vertebrates have a backbone, four appendages, paired kidneys and eyes, a single liver and heart, and so on. The anatomy of a tiny fish or lizard is very similar to that of humans. Chimpanzees and gorillas share our genes, proteins, and blood groups. The simplest explanation for such commonalities of form is shared descent, inheritance from a common ancestor. 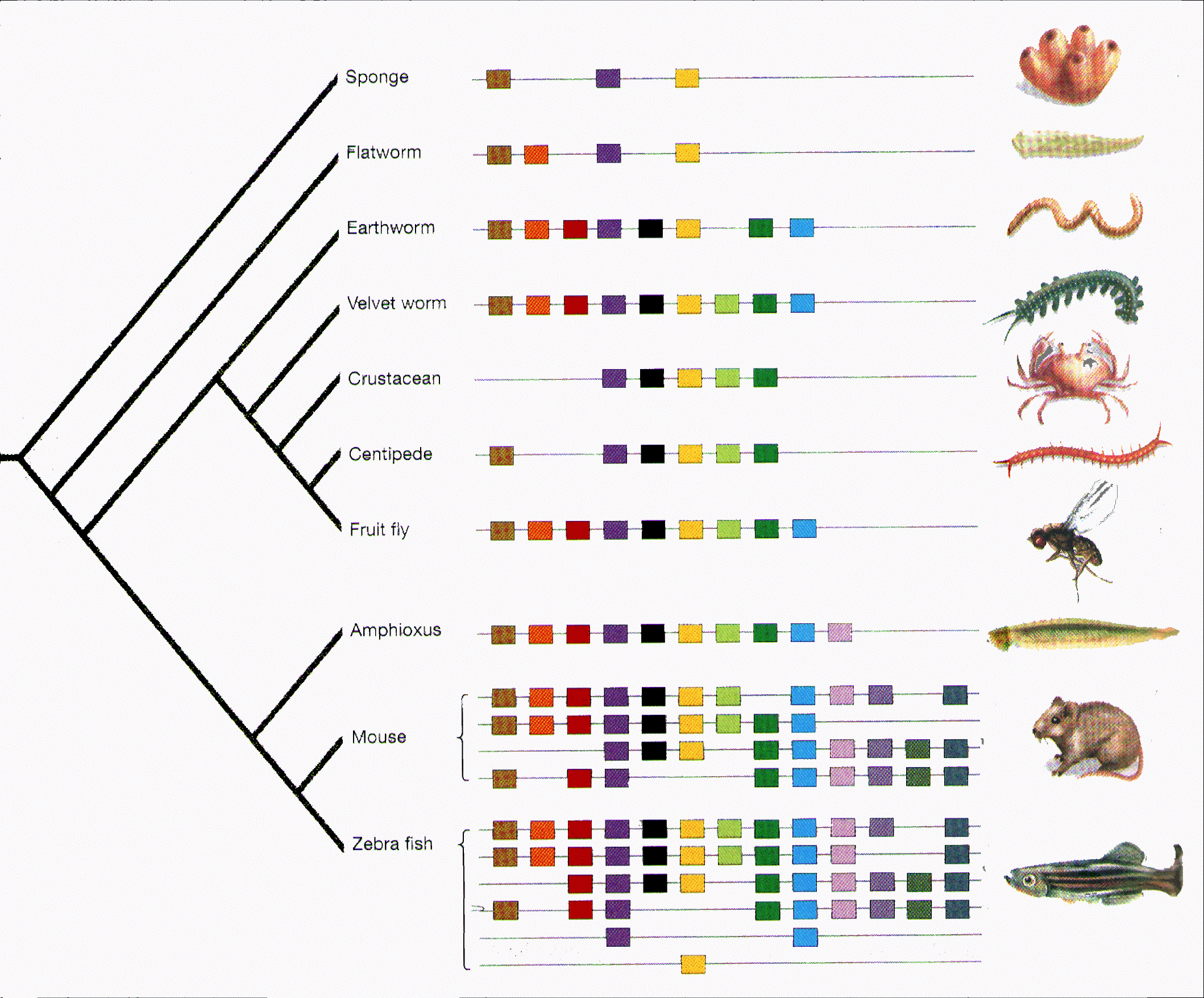
Invertebrates from sponges to flies share Hox genes with vertebrates. Many of the genes that drive our physiology and metabolic processes were actually invented billions of years ago by microbes in Earth's primeval oceans. Because they have no cell nuclei, bacteria are known as prokaryotes (pro = before, karyote = nucleus). Bacteria invented and perfected three fundamental metabolic processes: (1) photosynthesis, (2) fermentation, and (3) respiration. Photosynthesis is the process by which plants use light energy to convert inorganic carbon into energy-rich organic molecules. Oxygen is a byproduct of photosynthesis. Indeed, photosynthetic bacteria terraformed planet Earth, creating its oxygen atmosphere. Fermentation releases energy under anaerobic conditions, whereas respiration releases energy under aerobic conditions. All 3 processes are vital to life. In contrast, because "higher" organisms are more complex and have cell nuclei, they are known as eukaryotes (eu = true, karyote = nucleus). In addition to having a nucleus, eukaryotic cells house cell organelles known as mitochondria and chloroplasts, which are remnants of prokaryotes (see endosymbiosis. Mitochondria and chloroplasts (the latter are present only in plants) house a circular DNA structure homologous to those found in bacteria. These DNAs code for synthesis of enzymes essential to metabolism in all living things. Chloroplasts produce rubisco, a vital enzyme in the metabolic machinery responsible for photosynthesis, the process by which plants capture energy from light and convert it into chemical energy. Mitochondria are symbiotic cellular powerhouses housing ancient mitochondrial genes that first evolved billions of years ago and have been maintained by stabilizing natural selection ever since. Mitochondrial genes code for ribosomal RNAs and proteins such as dehydrogenase, oxidase, and ATPase enzymes. These enzymes drive both anaerobic and aerobic metabolic processes. Without these vital enzymes, eukaryotes could not exist. Bottom Line: Every cell in your body houses many mitochondria, which power your metabolism and keep you alive. Even our own blood plasma reflects our ancient origin: it is very close to ocean water. Terrestrial animals took the ocean with them when they emerged from the seas. We are but one small branch on the tree of life. We share most of our genes with all other organisms, including bacteria and fungi. Let us now consider some of the logical arguments and evidence in favor of evolution. Evolution is defined as any change in a gene pool. A gene pool is the collection of all the genes possessed by an interbreeding population of individual organisms. Genes are important because they are heritable, passed on from one generation to the next. Gene pools can be altered by several different agents including random genetic drift in small populations, gene flow (migration and interbreeding) between populations, mutation pressures, and meiotic drive (also known as segregation distortion). Natural Selection, defined as differential reproductive success, is another way in which gene pools can be changed. Natural selection is by far and away the most important evolutionary mechanism because it is the only one of these agents that results in adaptation, conformity between an organism and its environment. 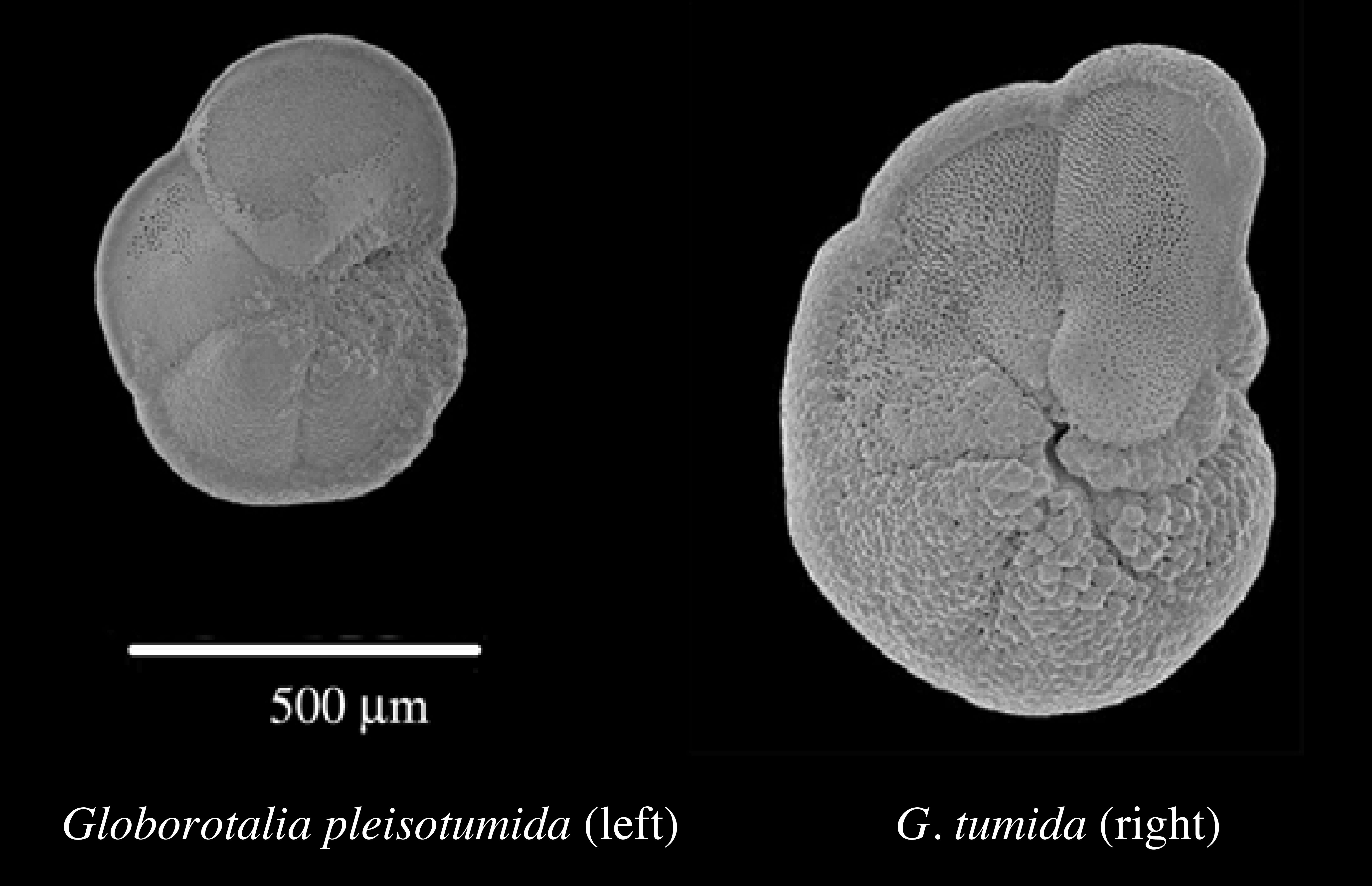
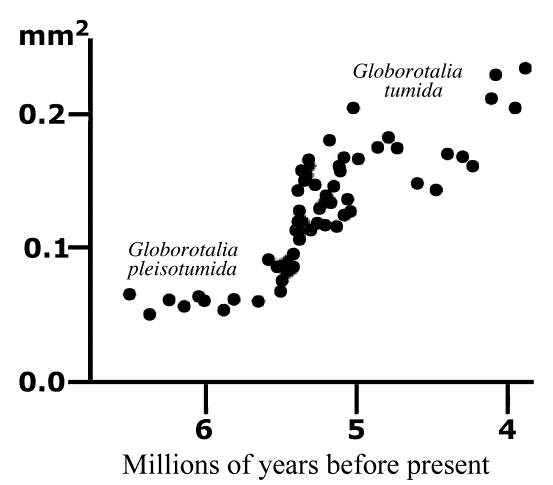
Evolutionary changes typically occur over long time periods, and fossil records are usually incomplete, making it difficult to document evolution. However, ocean sediments provide an uninterupted continuous record of fossil foraminiferans that increased in size over about a million years. These foraminifera also changed anatomically and are recognized as different species (see above figures). Humans have practiced anthropogenic natural selection or so-called ōartificial selectionö over many millenia in selectively breeding crop plants and the domestication of many animals. Biologists have observed evolution actually occurring over much shorter time scales in bacteria becoming resistant to antibiotics, insects evolving resistance to insecticides, stickleback fish adapting to changes in lake conditions, and the beaks of finches evolving in response to changing availability of seeds of different sizes. Anti-evolutionary creationists often assert that such examples are merely micro-evolution and that no evidence exists for macro-evolutionary changes in kind such as from a dog to a cat. Of course, dogs did not evolve into cats, but both share a common carnivorous ancestor that lived hundreds of millions of years earlier. The immense depth of time available for major evolutionary changes is difficult for humans to fathom. Incrementally slow changes over vast periods of time have resulted in spectacular changes in the body plans of plants and animals. 
Another fallacious argument anti-evolutionists often make is that ōsince evolution is random, how could it possibly lead to such complex designs?ö. The word ōrandomö is much abused term with varied meanings. The dictionary definition is ōwithout definite directionö or ōlacking a definite plan, purpose, or patternö or ōequi-probable.ö Often the word is used to describe anything capricious or unpredictable, which usually means that something is simply so poorly understood that it appears indeterminate. Invoking randomness may often merely be a cover up for our ignorance. Because the basic source of genetic variability is mutations caused by ōrandomö errors in copying DNA, creationists mistakenly assert that ōevolution is random.ö In actuality, of course, natural selection favors highly non-random organisms whose adaptations to cope with their environments enhance their reproductive success. Over eons of evolutionary time, natural selection has ferreted out detrimental mutations and favored those that are of benefit to their carriers. Now, letÆs consider complex networks of interacting species such as those that occur in natural ecological communities. Are these ōdesignedö for orderly and efficient function? Is there a natural ōbalance of nature?ö Although it may be tempting, it is dangerously misleading to view entire ecosystems as having been ōdesignedö for orderly and efficient function. Natural Selection simply does not operate for the ōgood of the speciesö but works by differential reproductive success of individual organisms. Antagonistic interactions at the level of individuals and populations are widespread: for example, these include competition, predation, and parasitism. Even the two parties engaged in a mutualism experience conflicts of interest because costs and benefits differ for each participant. Such negative interactions must frequently impair some aspects of ecosystem performance. Only a small fraction of the plant food on land is actually harvested by animals; most products of primary production are consumed by decomposers. Transfer of energy from one trophic level to the next higher trophic level is defined as ecological efficiency. Such efficiencies of transfer of energy from one trophic level to the next are low, generally only about 5 to 10 percent. Natural Selection operating on individual prey organisms favors escape ability, which in turn reduces the rate of flow of matter and energy through that trophic level, decreasing ecological efficiency but simultaneously increasing community stability. In contrast, predators evolve so as to be better able to capture their prey, which increases the efficiency of flow of energy through trophic levels but reduces a systemÆs stability. In the co-evolution of a predator and its prey, to avoid extinction, the prey must remain a step ahead of its predator. As a corollary, community-level properties of ecological efficiency and community stability may in fact be inversely related because natural selection operates at the level of individual predators and prey. Moreover, the apparent constancy and observed low levels of ecological efficiency are probably a result of this ōcompromiseö that must be reached between prey and their predators. Human attempts to increase ecological efficiency may well come at the cost of reduced stability. Because they want to think humans are devine, proponents of so-called ōintelligent designö (actually ardent creationists) are militantly opposed to the scientific theory of evolution by natural selection. By refusing to think rationally, these unfortunate and ignorant people have blinded themselves to the ability to understand the living world around them. See also: Anthropocentrism ERP: Natural Selection ERP: Convergent Evolution ERP: Intelligent Design? Noam Chomsky: Malignant Design? ERP: Arrogant Ignorance and Blind Optimism Reg Morrison's ōEvolution's Problem Gamblersö Jerry Coyne: Dead Genes and Vestigial Anatomy http://www.actionbioscience.org/evolution/nhmag.html |