Natural Selection © Eric R. Pianka Translated into Slovak by Barbara Lebedova Evolution is the gradual genetic change of living organisms over time due to ecological pressures they experience. Individuals in an interbreeding population share in a common gene pool. A population’s gene pool evolves, not an individual, due to changes occurring in such a gene pool. Evolution can occur in several different ways: (1) gene frequencies may be changed by migration, gene flow from another population. (2) in small populations, gene frequencies can be changed via random sampling -- this is known as genetic drift. (3) mutation pressure and (4) non-Mendelian segregation (meiotic drive) may also change gene pools. However, by far the most important agent of evolution is (5) natural selection, which operates by differential reproductive success of individuals. Natural selection is the only directed evolutionary mechanism resulting in conformity between an organism and its environment. This is how adaptations arise and are maintained. Natural selection is truly the fundamental unifying theory for all life. A thorough comprehension of natural selection opens a window of lucidity enabling an understanding of virtually any phenomenon in the living world -- it is a cosmic universal process, even for any non-DNA based alien life forms that may exist elsewhere in the Cosmos. As powerful as natural selection is, unfortunately, it is sometimes misunderstood. A persistent misconception is that natural selection occurs mainly through differences between organisms in death rates, or differential mortality (e.g. “Nature, red in tooth and claw”). 
Selection normally proceeds in a much more subtle and inconspicuous way. Whenever one organism leaves more successful offspring than another, in time its genes will come to dominate the population gene pool. Eventually, the genotype leaving fewer offspring must become extinct in a stable population, unless concomitant changes confer an advantage on it as it becomes scarcer. Ultimately, natural selection operates only by differential reproductive success. Differential mortality can be selective but only to the degree that it creates differences between individuals in the number of reproductive progeny they produce. Hence, phrases such as “the struggle for existence” and “survival of the fittest” have had an unfortunate consequence. They tend to make people think in terms of a dog-eat-dog world and to consider things such as predation and fighting over food as the prevalent means of selection. All too often, natural selection is couched in terms of differential death rates, with the strongest and fastest individuals considered as having a selective advantage over weaker and slower individuals. But, if this were the case, every species would continually gain in strength and speed. Because this is not happening, selection against increased strength and speed (counterselection) must be occurring and must limit the process. Animals can be too aggressive for their own good; an extremely aggressive individual may spend so much time and energy chasing other animals that it spends less than average time and energy on mating and reproduction, and as a result, leaves fewer offspring than average. Likewise, an individual can be too submissive and spend too much time and energy running away from other animals.  Differences in survivorship leading to differential mortality can, but need not always, lead to natural selection. A cautious tomcat that seldom crosses noisy streets may live to a ripe old age without leaving as many descendants as another less staid tom killed on a busy road at a much younger age.
Differences in survivorship leading to differential mortality can, but need not always, lead to natural selection. A cautious tomcat that seldom crosses noisy streets may live to a ripe old age without leaving as many descendants as another less staid tom killed on a busy road at a much younger age.  Unless living longer allows or results in higher reproductive success, long life is not favored by natural selection. Similarly, although we might wish otherwise, there is no necessary selective premium on beauty, brains, or brawn, unless such traits are in fact translated into more offspring than average. Thus, the only currency recognized by natural selection is babies. If ugly, dumb, weak, individuals leave more progeny, they will inherit the Earth. Natural selection operates like a short-sighted efficiency expert, ferreting out each and every way to maximize reproductive success, even when this becomes detrimental to survival. Selection is not sentient and has no ‘goal’ other than optimizing the use of resources towards immediate successful reproduction. Natural selection is also the ultimate inventor: a short list of its many patents includes flight, celestial navigation, many pharmaceuticals, echolocation, insulation, infrared sensors, and hypodermic needles. Unless living longer allows or results in higher reproductive success, long life is not favored by natural selection. Similarly, although we might wish otherwise, there is no necessary selective premium on beauty, brains, or brawn, unless such traits are in fact translated into more offspring than average. Thus, the only currency recognized by natural selection is babies. If ugly, dumb, weak, individuals leave more progeny, they will inherit the Earth. Natural selection operates like a short-sighted efficiency expert, ferreting out each and every way to maximize reproductive success, even when this becomes detrimental to survival. Selection is not sentient and has no ‘goal’ other than optimizing the use of resources towards immediate successful reproduction. Natural selection is also the ultimate inventor: a short list of its many patents includes flight, celestial navigation, many pharmaceuticals, echolocation, insulation, infrared sensors, and hypodermic needles.
 Natural selection has led to both instincts and learning. Learning is favored in unpredictable situations where animals can best adapt by altering their behavior appropriately. Instincts are basically "hard wired" learning and evolve under predictable situations, especially those that involve life and death decisions. Thus, because humans arose in Africa, surrounded by dangerous venomous snakes, we evolved an instinctive fear of snakes.
Natural selection has led to both instincts and learning. Learning is favored in unpredictable situations where animals can best adapt by altering their behavior appropriately. Instincts are basically "hard wired" learning and evolve under predictable situations, especially those that involve life and death decisions. Thus, because humans arose in Africa, surrounded by dangerous venomous snakes, we evolved an instinctive fear of snakes.
Many other emotions also evolved to adapt us to our environments.  For many milennia, humans lived as hunter gatherers and took refuge in caves. Greed and revenge must certainly have been adaptive for early cave dwellers. A greedy caveman who refused to share his food stores at the onset of winter would have been more likely to survive until spring and hence would have enjoyed higher fitness (reproductive success) than a non-greedy sharing caveman. Similarly, revenge made sense -- if another caveman messed with your stuff, you bashed him over the head and he was unlikely to do it again. Such instincts worked to our advantage when we were cavemen, but have become dangerously maladaptive in today's man-made artifical world. Currently, we worship runaway greed, allowing others to become billionaires -- what sense does it make to have more than you can actually use? Likewise, revenge makes no sense when one contemplates pushing a red button to set off nuclear explosives that will destroy yourself as well as your enemies.
For many milennia, humans lived as hunter gatherers and took refuge in caves. Greed and revenge must certainly have been adaptive for early cave dwellers. A greedy caveman who refused to share his food stores at the onset of winter would have been more likely to survive until spring and hence would have enjoyed higher fitness (reproductive success) than a non-greedy sharing caveman. Similarly, revenge made sense -- if another caveman messed with your stuff, you bashed him over the head and he was unlikely to do it again. Such instincts worked to our advantage when we were cavemen, but have become dangerously maladaptive in today's man-made artifical world. Currently, we worship runaway greed, allowing others to become billionaires -- what sense does it make to have more than you can actually use? Likewise, revenge makes no sense when one contemplates pushing a red button to set off nuclear explosives that will destroy yourself as well as your enemies.
Hence, natural selection can backfire, as it has many times. For example, selection has programmed us to reproduce by making our nerve endings tingle in the right places -- cave people made plenty of babies without even knowing where they came from!  Like all life forms, we are mere puppets dancing to the strings of natural selection, often without even being aware of it. Humans have overpopulated
Spaceship Earth
and we are destroying our own life support systems, actually threatening to destroy ourselves. Natural selection does not often favor "niceness" but usually leads to selfish behaviors. Even in cooperation, such as mutualisms, parties differ in costs and benefits gained. For such cooperative systems to evolve, net benefits to both parties must outweigh costs. Unfortunately, as powerful as natural selection is, it is extremely short sighted, and thus an imperfect, efficiency expert. You might even argue that natural selection is our biggest enemy -- to live in a sustainable long-term equilibrium with Earth's finite resources, humans will have to overcome their preprogrammed urge to reproduce. That won't be easy and may even be impossible.
Like all life forms, we are mere puppets dancing to the strings of natural selection, often without even being aware of it. Humans have overpopulated
Spaceship Earth
and we are destroying our own life support systems, actually threatening to destroy ourselves. Natural selection does not often favor "niceness" but usually leads to selfish behaviors. Even in cooperation, such as mutualisms, parties differ in costs and benefits gained. For such cooperative systems to evolve, net benefits to both parties must outweigh costs. Unfortunately, as powerful as natural selection is, it is extremely short sighted, and thus an imperfect, efficiency expert. You might even argue that natural selection is our biggest enemy -- to live in a sustainable long-term equilibrium with Earth's finite resources, humans will have to overcome their preprogrammed urge to reproduce. That won't be easy and may even be impossible.
Naiive, group selectionist explanations are often given for biological phenomena. For example, people often talk about things that "are good for the species." Natural selection simply does not work at that level, but instead favors reproductive success of individual organisms. Thinking about the evolution of various ecological attributes must be done carefully and correctly. For example, it is tempting, but dangerously misleading, to view organisms or ecosystems as having been “designed” for orderly and efficient function. Numerous attributes of individuals are poorly designed carryovers from ancestors with different ecologies. Adapting an ancestral fish into a land dwelling mammal necessarily involved many changes of function and led to some elements of poor design, such as the crossover between respiratory and ingestion tubes resulting in a maladaptive lung/esophagus arrangement (one that leads to almost 3,000 choking deaths in America each year). One must always keep in mind that natural selection operates by differential reproductive success of individual organisms. Antagonistic interactions at the level of individuals and populations (competition, predation, parasitism) must frequently impair some aspects of ecosystem performance. Two fundamentally different approaches have proven useful in biology, the functional (mechanistic) and the evolutionary (strategic) approach. The former deals with a shorter time scale than the latter. These have been called the “how?” versus “why?” questions. For example, one might ask “why are sex ratios often near 50:50?” A proximate mechanistic answer might be that (in mammals, at least) half the sperm contain a y-chromosome, while the other half contain the x chromosome: thus half the zygotes formed will be xy (male) and the other half will be female (xx).  Fisher (1930) offered an evolutionary answer: no matter what the current sex ratio, each and every generation, half the genes in the next generation’s gene pool must come from males and the other half from females. Thus, investing in sons has the same expectation of payoff as investing in daughters: at equilibrium, parents should invest equally in the two sexes, which will usually result in a sex ratio near 50:50. Fisher went on to note that, in humans, sex ratios at birth are actually statistically biased towards an excess of males. Then, connecting the dots, he noted that infant mortality is higher among human males than females. Hence, to equalize parental expenditure on sons and daughters, parents must over invest in sons early in the period of parental care and compensate for this later during parental care by investing more in daughters. Fisher concludes “since this adjustment is brought about by a somewhat large inequality in the sex ratio at conception, for which no a priori reason can be given, it is difficult to avoid the conclusion that the sex ratio has really been adjusted by these means.”
Fisher (1930) offered an evolutionary answer: no matter what the current sex ratio, each and every generation, half the genes in the next generation’s gene pool must come from males and the other half from females. Thus, investing in sons has the same expectation of payoff as investing in daughters: at equilibrium, parents should invest equally in the two sexes, which will usually result in a sex ratio near 50:50. Fisher went on to note that, in humans, sex ratios at birth are actually statistically biased towards an excess of males. Then, connecting the dots, he noted that infant mortality is higher among human males than females. Hence, to equalize parental expenditure on sons and daughters, parents must over invest in sons early in the period of parental care and compensate for this later during parental care by investing more in daughters. Fisher concludes “since this adjustment is brought about by a somewhat large inequality in the sex ratio at conception, for which no a priori reason can be given, it is difficult to avoid the conclusion that the sex ratio has really been adjusted by these means.”
For many years, biologists merely accepted a broad range of biological phenomena as essentially immutable, such as the above example that sex ratios are usually near equality, without considering why such facts might be so or how they could have evolved. Up until about 1970, an evolutionary perspective was totally lacking in ecology textbooks. An intellectual revolution has taken place in ecology during the past half century: rigorous application of the theory of natural selection in population biology has greatly increased our understanding of numerous phenomena, including the evolution of such things as genetic dominance, foraging modes, reproductive tactics, senescence, menopause, sex ratios, tolerance curves, a wide range of social behaviors, mate choice, mating systems, predator escape tactics, parasite virulence, host-altered behavior, evolutionary epidemiology, Darwinian medicine, niche breadth, guild structure, and resource partitioning, among others. The addition of an evolutionary approach has opened up whole new areas of endeavor such as optimal foraging, life history tactics, sexual selection, and coevolution, each of which has become an instant subdiscipline and then quickly exploded into a field in its own right. Evolutionary ecology has blossomed into a massive discipline that has assimilated and largely replaced other ecological subdisciplines. Modern molecular techniques now allow biologists to isolate, amplify and sequence DNA, which in turn can be used to reconstruct probable evolutionary trees and ancestral states. Phylogeny and modern comparative methods now allow ecologists to deduce and trace the probable actual course of evolution. 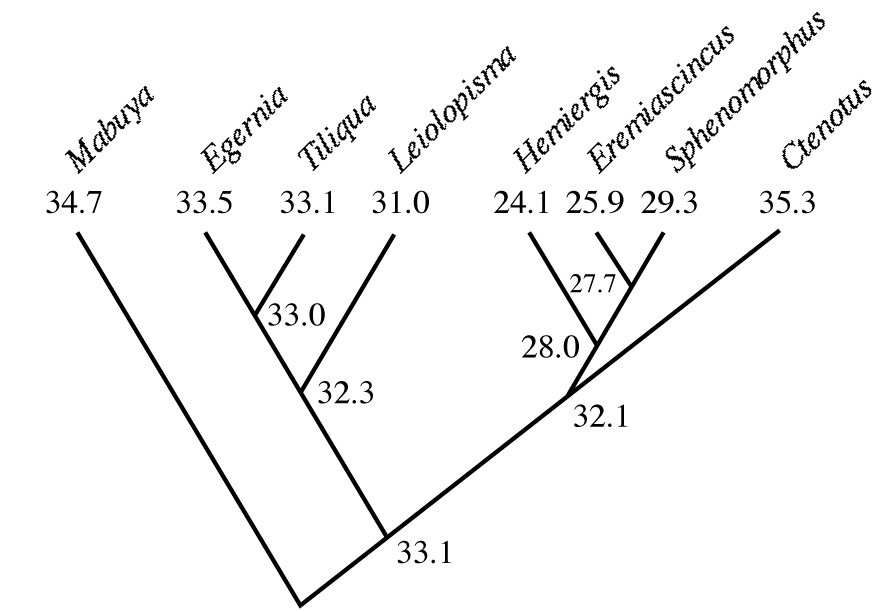
Active body temperatures in °C among seven genera of Australian skinks, with inferred ancestral body temperatures at various nodes. Note the high body temperature of Ctenotus (far right) and lower body temperatures of its relatives Hemiergis, Eremiascincus, and Sphenomorphus. This phylogenetic analysis suggests that descendent lineages diverged from a common ancestor, which possessed a moderate active body temperature [Adapted from Huey and Bennett (1987).] All life on Earth arose from one common ancestor whose descendents underwent divergent selection similar to that shown in the above figure.  Darwin ended “The Origin of Species” with “It is interesting to contemplate a tangled bank,
clothed with many plants of many kinds, with birds singing on the bushes, with various
insects flitting about, and with worms crawling through the damp earth, and to reflect that
these elaborately constructed forms, so different from each other, and dependent upon each
other in so complex a manner, have all been produced by laws acting around us. These laws,
taken in the largest sense, being growth with reproduction; inheritance which is almost
implied by reproduction; variability from the indirect and direct action of the conditions
of life, and from use and disuse; a ratio of increase so high as to lead to a struggle for
life, and as a consequence to natural selection, entailing divergence of character and the
extinction of less improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production of the higher
animals, directly follows. There is grandeur in this view of life . . .”
Darwin ended “The Origin of Species” with “It is interesting to contemplate a tangled bank,
clothed with many plants of many kinds, with birds singing on the bushes, with various
insects flitting about, and with worms crawling through the damp earth, and to reflect that
these elaborately constructed forms, so different from each other, and dependent upon each
other in so complex a manner, have all been produced by laws acting around us. These laws,
taken in the largest sense, being growth with reproduction; inheritance which is almost
implied by reproduction; variability from the indirect and direct action of the conditions
of life, and from use and disuse; a ratio of increase so high as to lead to a struggle for
life, and as a consequence to natural selection, entailing divergence of character and the
extinction of less improved forms. Thus, from the war of nature, from famine and death, the
most exalted object which we are capable of conceiving, namely, the production of the higher
animals, directly follows. There is grandeur in this view of life . . .”
For the first and only time in the entire 3.5+ billion year history of life on Earth, a product of natural selection has evolved intelligence enough to become aware of the force (natural selection) driving all living systems.  Essentially, to
paraphrase Balfour (1895) "
matter knows itself."
We have the ability to understand the evolutionary basis of human emotions: betrayal, compassion,
empathy, envy, fear, greed, honesty, jealousy, loyalty, lust, revenge, and trust. But we still
lack the ability to control them. Unfortunately, rather than use our intelligence to become
god-like stewards of this planet, humans have instead chosen to rape and destroy the only Earth
we have. Essentially, to
paraphrase Balfour (1895) "
matter knows itself."
We have the ability to understand the evolutionary basis of human emotions: betrayal, compassion,
empathy, envy, fear, greed, honesty, jealousy, loyalty, lust, revenge, and trust. But we still
lack the ability to control them. Unfortunately, rather than use our intelligence to become
god-like stewards of this planet, humans have instead chosen to rape and destroy the only Earth
we have.
People now dominate Earth’s ecosystems to such an extent, pure ecology has all but vanished from the face of this planet! Pristine ecological systems no longer exist anywhere. Multitudinous anthropogenic effects of overpopulation include many different kinds of pollution of the atmosphere, water, and land (and the manifold effects of such pollution on the health and livelihood of plants and animals, including ourselves), habitat destruction and fragmentation, endangered species, loss of genetic variability, extinction, disruption of natural ecosystems, human transportation of organisms and the resultant homogenization of earth’s biota, evolution of microbes that infect humans as hosts, and even murder rates among humans. Until recently, spaceship Earth  has provided us with a rather nice place to live. But now, Earth’s life support systems are
failing . . . we have overpopulated the planet and fouled its atmo-sphere -- the resultant
pollution is contributing to global weather change.
has provided us with a rather nice place to live. But now, Earth’s life support systems are
failing . . . we have overpopulated the planet and fouled its atmo-sphere -- the resultant
pollution is contributing to global weather change.
 Earth is
warming rapidly-- ice caps are melting and ocean currents are changing.
Polar bears
and penguins are facing extinction and though many humans refuse to face the facts, we might not be far behind. Earth is
warming rapidly-- ice caps are melting and ocean currents are changing.
Polar bears
and penguins are facing extinction and though many humans refuse to face the facts, we might not be far behind.
See also: ERP: Evolution ERP: Adaptation ERP: Coevolution ERP: Convergent Evolution ERP: Reproductive Tactics ERP: Sexual Selection ERP: Scientific Methods and Human Knowledge References Balfour, AJ (1895) Foundations of Belief. Longmans Green. Darwin C (1859) The origin of species by means of natural selection. Murray, London. Fisher RA. (1930) The genetical theory of natural selection. Clarendon Press, Oxford. Garland T, RB Huey, & AF Bennett (1991) Phylogeny and coadaptation of thermal physiology in lizards: A reanalysis. Evolution 45: 1969-1975. Huey RB & AF Bennett (1987) Phylogenetic studies of coadaptation: Preferred temperatures versus optimal performance temperatures of lizards. Evolution 41: 1098-1115. Hutchinson GE (1959) Homage to Santa Rosalia, or why are there so many kinds of animals? Amer. Natur. 93: 145-159. Pianka, ER (2000) Evolutionary Ecology, 6th ed. Addison-Wesley Longman. To go to Pianka Lab Homepage |