1| Background
Humans seem to delight in animal motifs -- thus, we have automobiles, airplanes, and athletic teams named after various animals: cougar, jaguar, lynx, mustang, pinto, ram, eagle, falcon, nighthawk, roadrunner, and the list goes on. Zoos are a popular form of entertainment, particularly for children. Yet, many people feel threatened by a free-ranging wild creature, even by a tiny mouse or a harmless snake. Indeed, urbanization is now so complete that, aside from cockroaches and songbirds (and perhaps while on vacation), most of us seldom encounter wild animals.
What is the essential difference between a wild animal versus one in a cage? Clearly, a rattlesnake behind glass does not pose nearly as much physical danger to a human observer as does a wild rattlesnake. For the study of many kinds of biological phenomena, there is in fact no difference between a caged specimen, so long as it remains alive, and its wild cousin. The constrained one still has intact cells, molecules, physiological processes, and, to some extent, behavior. But the caged animal, removed from its habitat, is out of context -- it has been stripped of its natural history and it no longer interfaces with the environment to which it is adapted and in which it evolved.
The biological discipline of ecology deals with a myriad of ways in which organisms (plants, animals, and other heterotrophs such as bacteria and fungi) interact with, influence, and are in turn influenced by their natural surroundings. Wild plants and animals in their natural communities constitute the subject matter of ecologists. To these scientists, caged organisms might as well be dead for they have no ecology. Ecology differs from other sorts of biology in that its perspective is directed upward and outward from the individual organism to its environment. Other kinds of biology focus on organismal and suborganismal processes and thus involve a reductionistic viewpoint. Ecology has a more holistic perspective.
Scaling in Biology
Biology is a vast discipline ranging from molecules to cells to tissues to whole organisms to kin groups to
populations to communities (and clades) to entire ecosystems, or even the entire global biosphere. Across this broad range of scale,
factors vary by many orders of magnitude (Figure 1.1).
-
Figure 1.1. Diagrammatic representation of the time-space scaling of various biological phenomena. Community and ecosystem phenomena occur over longer time spans and more vast areas than suborganismal- and organismal-level processes and entities. [Redrawn from Osmund et al. (1981)]
Molecular biology can be done quickly in small spaces, but community ecology requires decades and square kilometers. Biogeographic and historic events take place over many millennia. Continental plates have moved across thousands of kilometers over geological time. Until recently, ecologists have been preoccupied with local phenomena and events occurring on relatively short time scales. However, emerging new subdisciplines of landscape ecology and macroecology offer promising new regional and global perspectives.
As an example of the effects of scale, consider movements of organisms across landscapes (Brooks 1988). At the shortest temporal and spatial scale, individual organisms move during their lifetimes and disperse; over an intermediate scale in space and time, immigration and emigration occur between and among populations (see also section on metapopulations in Chapter 19); over a much greater spatial extent and at a much longer time scale, geographical ranges shift in response to climatic changes and to vicariant events such as geotectonic movements, leading ultimately to the formation of geographical patterns in species diversity.
Scientists are motivated by curiosity about their surroundings; they go to great lengths to satisfy their desire to understand natural events and phenomena. All scientists assume that an organized reality exists in nature and that objective principles can be formulated that will adequately reflect this natural order. A fundamental and important way in which biological phenomena can be ordered is by simple and direct enumeration, as in the classification of organisms or biotic communities. Thus, ecologists recognize different ecological systems such as tundra, desert, prairie, savanna, deciduous forest, coniferous forest, and rain forest. Early ecology was primarily descriptive; the originators of the science spent most of their time describing, itemizing, and classifying various ecological elements. This process was absolutely essential before a more process-oriented ecology could be developed.
Founded and firmly based on this older body of descriptive information, modern ecology seeks to develop general theories with predictive powers that can be compared against the real world. Ecologists want to understand and to explain, in general terms, the origin and mechanisms of interactions of organisms with one another and with the nonliving world. To build such general theories of nature, ecologists construct hypotheses, hypothetical "models" of reality. All models must make simplifying assumptions -- some sacrifice precision for generality, whereas others sacrifice generality for precision. Some models actually sacrifice certain aspects of realism itself! Models have been described as "mere caricatures of nature designed to convey the essence of nature with great economy of detail." No model is "correct" or "true" -- any given model merely represents one particular attempt to mimic reality. All models are to some extent incorrect. To be most useful, models are usually designed to generate testable predictions. Most models can therefore be confronted with reality and can be falsified. But not all models are refutable; some conceptual models have proven to be useful in an abstract way even though they do not suggest direct tests.
When a model's predictive powers fail, it is either discarded or revised (Popper 1959). Models and hypotheses that do not conform adequately to reality are gradually replaced by those that better reflect the real world (Lakotos 1970). The scientific method is thus self-regulating; as time progresses, knowledge expands and is continually refined and improved to reflect external reality better and better. Well-substantiated hypotheses become theories. We are very fortunate indeed to be able to benefit from past genius and research effort; in a few hours of careful reading, you can now learn material that required many lifetimes to acquire.
Due to the multiple meanings of words, verbal models are usually somewhat ambiguous and imprecise and therefore of limited utility. The great complexity of ecological systems necessitates the use of graphical and mathematical models, so much so that ecologists often employ nearly as much mathematics as biology. However, the development of sound ecological principles depends heavily on what might be called "biological intuition," and there is certainly no substitute for a firm foundation in natural history. Models based on erroneous biological assumptions, no matter how elegant and elaborate, can hardly be expected to reflect nature accurately. Hence, a good background for comprehending ecology includes some biology and mathematics as well as a solid basis in general science.
Scientists are often said to be working with facts, and human knowledge is supposed to be based on them. But what exactly is a "fact" ? The dictionary definition is "what has really happened or is the case; truth; reality." Consider, for example, the apparently simple fact that the sun rises each morning. Daily we receive new evidence confirming this fact. We are quite confident that the sun will rise again tomorrow, but there is a remote possibility that someday it may not. Indeed, on a time scale of eternity, we can be certain that someday it will burn itself out and conjecture that eventually it will cease to exist. Appearances to the contrary, even simple facts such as the sunrise are not "clean" of interpretation (Nietzsche wrote "there are no facts, only interpretations"). Under the now defunct "flat earth-moving sun" hypothesis, the sun's movement across the sky was viewed from the perspective of a fixed nonmoving earth at the center of the universe. Indeed, references to sunrise and sunset are based on this interpretation, which is supported by our superficial commonsense perceptions. But another hypothesis has supplanted the concept of a moving sun; our understanding of cosmic events is greatly enhanced when the sun is viewed as fixed in space at the center of a solar system, and the earth is interpreted as a rotating globe orbiting around a small star (Copernicus). Our vocabulary hasn't caught up -- we should refer to "sunrise" and "sunset" as "Earthspin Dawn" and "Earthspin Dusk." Perhaps someday another even more powerful model will replace our current working hypothesis "explaining" sunrise.
Observation and experiment play an important and vital role in science. They are used to test models, to refute inadequate hypotheses, and thus they help us to formulate improved interpretations of natural phenomena. Certain kinds of natural events cannot be manipulated. Thus, we cannot stop the sun's fusion or earth's rotation to test our current ideas, but each daily observation of sunrise or sunset nevertheless strengthens our confidence in the accepted interpretation of celestial events. Similarly, in many ecological situations (particularly those involving the evolution of adaptations), direct experimentation is often impractical or even impossible if not illegal or immoral. Some sorts of ecological phenomena do lend themselves to manipulation, but like astronomers, evolutionary ecologists rely heavily on a careful comparative approach (sometimes referred to as a "natural" experiment).
A major goal of science is to understand causality; when we observe that event B always follows event A, we sometimes infer that A causes B. But some other unknown event X could be causing both to occur with a time lag in B. The way scientists test for such spurious correlations is to find a way to cause event A to occur, holding as much else as possible constant in a controlled experiment; then, if B still follows A when X does not occur, we are more confident that A indeed causes B. In the physical sciences and in mechanistic kinds of biology such as molecular biology, causal connections are usually fairly simple and straightforward, allowing construction of mutually exclusive alternative hypotheses (Platt 1964). Scientific investigation can then proceed by what has been termed "strong inference," and researchers can choose among logical alternatives. But in population ecology, mutually exclusive alternatives often cannot be formulated. Multiple causality is common and ecologists can seldom eliminate a hypothesis as inadequate under all situations.
Domain of Ecology: Definitions and Groundwork
Ecology has been variously defined as "scientific natural history," "quantitative natural history," "the study of the structure and function of nature," "the sociology and economics of animals," "bionomics," "the study of the distribution and abundance of organisms," and "the study of the interrelationships between organisms and their environments." The last of these definitions is probably the most useful, with " environment" being defined as the sum total of all physical and biological factors impinging on a particular organismic unit.
For "organismic unit," one can substitute either "individual," "family group," "population," "species," or "community." Thus, we may speak of the environment of an individual or the environment of a population, but to be precise, a particular organismic unit should be understood or specified. The environment of an individual contains fewer elements than the environment of a population, which in turn is a subset of the environment of the species or community.
To avoid the apparent circularity in the preceding definition, ecology might be better defined as the study of the relations between organisms and the totality of the physical and biological factors affecting them or influenced by them. Thus, ecologists begin with the living organism and seek to understand how an organism affects its surroundings and how those surroundings in turn affect the organism.
Environment includes everything from sunlight and rain to soils and other organisms. An organism's environment consists not only of other plants and animals encountered directly (such as foods, trees used for nesting sites, potential predators, and possible competitors), but also of purely physical processes and inorganic substances, such as daily temperature fluctuations and oxygen and carbon dioxide concentrations. Of course, the latter may be affected by other organisms, which then indirectly become a part of the environment of the first organism. Indeed, any remote connection or interaction between two organismic units dictates that each is a part of the other's environment.
Pristine natural environments (those that remain relatively unaltered by humans) are of particular interest and importance in evolutionary ecology because they constitute the environments to which any particular organism has become adapted over evolutionary time. Because there are direct or indirect interactions between almost all the organisms in a given area, the biotic component of the environment of most organisms is extremely complex. Coupling this great complexity with a multifaceted and dynamic physical environment makes ecology an exceedingly vast subject. No other discipline seeks to explain such a wide variety of phenomena at so many different levels. As a consequence, ecology takes in aspects of numerous other fields, including physics, chemistry, mathematics, computer science, geography, climatology, geology, oceanography, economics, sociology, psychology, and anthropology. Ecology is properly classified as a branch of biology; students of ecology attempt to interweave and correlate the subdisciplines of biology such as evolution, genetics, systematics, morphology, physiology, ethology (behavior), as well as various taxonomic subdivisions of biology like algology, entomology, ichthyology, herpetology, mammalogy, and ornithology. Sometimes "plant ecology" is distinguished from "animal ecology." As basic as this distinction may be, however, it is most unfortunate; plants and animals inevitably constitute important parts of one another's environments, and their ecologies simply cannot be disentangled. Interactions between plants and animals have recently been the subject of considerable thought and field study.
Obviously no one can master all of such an enormous field, and as a result, there are many different kinds of ecologists with a wide variety of expertise and perspectives on the subject matter of the science. The breadth and difficulty of ecology, combined with its youth and great relevance to urgent human problems, makes it a fascinating and exciting field. Major basic discoveries of real importance remain to be made, and the potential for growth and refinement is immense. Young sciences, and especially complex biological ones like ecology, have sometimes been characterized as "soft" sciences, because they are not as precise as the older and better established "hard" sciences such as chemistry and physics. Of course, by its very nature, the subject matter of biology is innately much more diverse and complex, not to mention interesting, than that of physics and chemistry. As every science matures, it becomes more and more abstract and its hypotheses are refined, tested, and improved until they eventually attain the status of theories or "laws," as in the familiar laws of chemistry and physics. Ecology at present has few firm laws but many hypotheses, and much work on and testing of these hypotheses remains to be done. The single concept closest to deserving the status of "law" in ecology and one that is shared with all of biology, is natural selection (see section below).
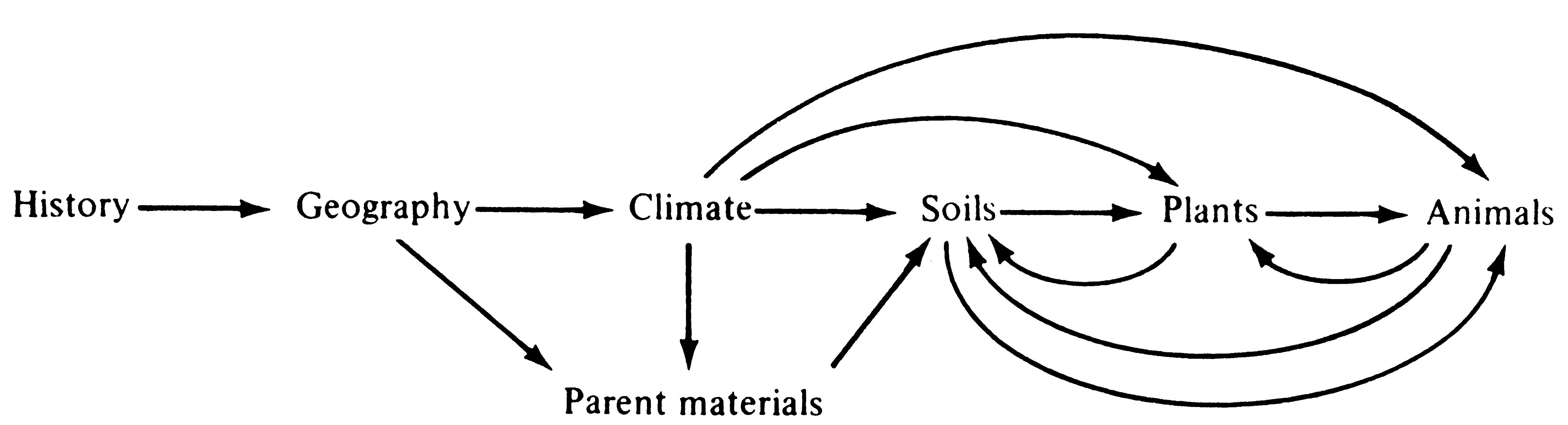
Ecology encompasses many complexly interrelated concepts and involves phenomena at several different levels of organization. Indeed, the subject matter of the discipline is inherently multidimensional. But language forces us to express ourselves in a one-dimensional stream of words. How, then, will we unravel this unfinished tapestry that has no beginning and no end? Components of ecological systems can be conveniently considered in order of generally increasing complexity, proceeding from the inorganic to the organic world (Figure 1.2) and then from individuals to populations to communities. In this book, the organismic world is treated in much greater detail than the non-organismic world.
-
Figure 1.2. Diagrammatic representation of the subject matter of ecology showing the natural sequence proceeding from the inorganic to the organic world. Many other arrows and feedback loops could be added, but those depicted are of major importance. Chapter 2 deals with history; Chapters 3 and 4 treat geography, climate, soils, and plants. Chapter 5 examines how organisms cope with their physical environments. Remaining chapters look at the interactions between and among plants and animals, especially at the population and community levels.
The climate, soils, bacteria, fungi, plants, and animals at any particular place together constitute an ecosystem. Ecosystems have both abiotic (nonliving) and biotic (living) components. All of the biotic components of an ecosystem, or all the organisms living in it, taken together, comprise an ecological community. The abiotic components can be separated into inorganic and organic, whereas the biotic components are usually classified as producers, consumers, and decomposers. Producers, sometimes called autotrophs (= self feeding), are the green plants that trap solar energy and incorporate it into energy-rich chemicals such as glucose. Consumers, or heterotrophs, are all the organisms (bacteria, fungi, and animals) that either eat the plants or one another; all heterotrophs thus depend, directly or indirectly, on plants for energy. Several levels of consumers are recognized (primary, secondary, and tertiary), depending on whether they eat plants directly or instead consume other herbivorous or carnivorous animals. Decomposers, which are also heterotrophs, are often bacteria and fungi; they break down plant and animal material into simpler components and thereby return nutrients to the nutrient pool and autotrophs. Decomposers are therefore essential in recycling matter within an ecosystem.
As previously indicated, plants and animals in ecosystems can be considered at several different levels: individuals, family groups, populations, species, and communities (subsequent chapters deal with each organismic level of organization, but most emphasis is given to populations). None of these levels of organization can be adequately understood in isolation because each exerts strong influences upon the others. Every individual is simultaneously a member of a population, a species, and a community, and it must therefore be adapted to cope with each and must be considered in that context. An individual's fitness -- its ability to perpetuate itself as measured by its reproductive success -- is determined not only by its status within its own population but also by the various interspecific associations of its species and especially by the particular community in which it exists. Similarly, every community is composed of many populations and numerous individuals, which determine many, but by no means all, of its properties. At each level of organization, important new properties emerge that are not properties of the preceding level. Thus, individuals have a fixed genetic makeup and live or die, whereas populations have gene frequencies, birth rates, and death rates. All these population parameters (and others) can change in time as the composition of a population changes in response to a changing environment ( evolution).
The Urgency of Basic Ecological Research
In one sense, ecology is the oldest and most basic science. Primitive humans simply had to have been astute natural historians to survive. As a more rigorous scientific discipline, however, ecology promises to be the shortest-lived science of all time. The word itself seems to have been first used in Germany in the late 1860s by E. Haeckel, but did not come into common use until 1895, when J. Warming used it in a book title. The first scientific society, the British Ecological Society, was founded in 1913. The earliest textbooks were published in 1927 (Elton) and 1929 (Weaver and Clements). Ecology is thus barely a century old and would seem doomed not to celebrate much of a bicentennial, for by then most natural ecological systems will surely be long gone. Although this realization is saddening, this "doomsday ecologist" nevertheless urges everyone to learn as much ecology as he or she can if for no reason other than simply because very soon we will need all of it that we can possibly get.
Ecology and environment are words frequently encountered in the news and popular media, almost invariably bandied about in conjunction with humans and their environment. As often as not the terms are misused, especially by politicians and other advertisers. Many people now use "ecology" to refer primarily to applied and human ecology. "Ecologist" has been equated with "rabid environmentalist" (as a result, many real ecologists now refer to themselves as population biologists!). The basic science of ecology is not synonymous with a study of the effects of people on their own surroundings and on other organisms, but in fact represents a much broader and more fundamental class of subject matter. Some problems facing humans today illustrate what can happen when ecological systems are not used wisely in accordance with sound ecological principles; as such, the content of this book is very pertinent to human ecology. However, throughout the book, emphasis is given to principles of basic ecology, particularly as these principles apply to and can be interpreted in terms of the theory of natural selection. Major concepts and principles are stressed more than detail, but references are given at the end of each chapter for those desiring to delve more deeply into particular subjects.
Most people consider the study of biology, particularly ecology, to be a luxury that they can do without. In the interests of obtaining "well-rounded" students, many medical schools no longer even require that premedical students major in biology. The study of basic biology is not a luxury at all, but rather an absolute necessity for living creatures. Despite our anthropocentric (human-oriented) attitudes, other life forms are not irrelevant to our own existence. For example, an understanding of basic parasitology is needed to control epidemics in human populations. Similarly, a knowledge of basic principles of community organization and ecosystem function is essential for wise exploitation of both natural and agricultural ecological systems. Beyond such anthropocentric arguments, one can argue that other species have a right to exist, too, as products of natural selection that have adapted to natural environments over millennia. With human populations burgeoning and pressures on space and other limited resources intensifying, we will need all the biological knowledge that we can get. Ecological understanding will prove to be particularly vital.
There is a great urgency to basic ecological research simply because the worldwide press of humanity is rapidly driving other species to extinction and destroying the very systems that ecologists seek to understand. No natural community remains pristine. Unfortunately, many will disappear without even being adequately described, let alone remotely understood. As existing species go extinct and even entire ecosystems disappear, we lose forever the very opportunity to study them. Knowledge of their evolutionary history and adaptations vanishes with them: we are thus losing access to biological information itself. Indeed, "destroying species is like tearing pages out of an unread book, written in a language humans hardly know how to read" (Rolston 1985). Just as ecologists are finally beginning to learn to read this "unread" and rapidly disappearing book of life, they are encountering governmental and public hostility and having a difficult time attracting support. This is simply pitiful. And time is quickly running out.
Darwin's theory of natural selection is a
truly fundamental unifying theory of life.
A thorough appreciation of the process of
selection is essential background to understanding evolutionary ecology. Natural
selection comes as close to being a "fact"
as anything in biology. Although there is
no such thing as "proof" in science (except
in mathematics, where all postulates are
taken as given), over the past century an
enormous body of data has been amassed in
support of the theory of natural selection.
Natural selection is not a difficult concept, but it is frequently misunderstood.
 Charles Darwin Charles Darwin
Natural selection is not synonymous with evolution. Evolution refers to temporal changes of any kind, whereas natural selection specifies one particular way in which these changes are brought about. Natural selection is the most important agent of evolutionary change simply because it results in conformity between organisms and their environments, or adaptations. Other possible mechanisms of evolution besides natural selection include inheritance of acquired characteristics, gene flow, meiotic drive, and genetic drift. Another persistent misconception is that natural selection occurs mainly through differences between organisms in death rates, or differential mortality.
Selection may proceed in a much more subtle and inconspicuous way. Whenever one organism leaves more successful offspring than another, in time its genes will come to dominate the population gene pool. Eventually, the genotype leaving fewer offspring must become extinct in a stable population, unless there are concomitant changes conferring an advantage on it as it becomes rarer. Ultimately, natural selection operates only by differential reproductive success. Differential mortality can be selective but only to the degree that it creates differences between individuals in the number of reproductive progeny they produce. Hence, phrases such as "the struggle for existence" and "survival of the fittest" have had a rather unfortunate consequence. They tend to make people think in terms of a dog-eat-dog world and to consider things such as predation and fighting over food as the prevalent means of selection. All too often, natural selection is couched in terms of differential death rates, with the strongest and fastest individuals considered to have a selective advantage over weaker and slower individuals. But if this were the case, every species would continually gain in strength and speed. Because this is not happening, selection against increased strength and speed (counterselection) must be occurring and must limit the process. Animals can sometimes be too aggressive for their own good; an extremely aggressive individual may spend so much time and energy chasing other animals that it spends less than average time and energy on mating and reproduction, and as a result, leaves fewer offspring than average. Likewise, an individual can be too submissive and spend too much time and energy running away from other animals.
Differences in survivorship leading to differential mortality can, but need not always, lead to natural selection. A cautious tomcat that seldom crosses noisy streets may live to a ripe old age without leaving as many descendants as another less staid tom killed on a busy road at a much younger age. Unless living longer allows or results in higher reproductive success, long life cannot be favored by natural selection. Similarly, although we might wish otherwise, there is no necessary selective premium on beauty, brains, or brawn, unless such traits are in fact translated into more offspring than average.
Some of the best documented examples of natural selection in action concern pesticide resistance in certain insects and drug resistance in many microbes. Other examples include industrial melanism in moths and the evolution of myxoma virus and Oryctolagus rabbits in Australia (for discussion, see Chapter 15).
Natural selection is the ultimate inventor: a short list of its many patents includes flight, celestial navigation, echolocation, insulation, infrared sensors, and hypodermic needles.
Some words of warning are now appropriate. Overenthusiastic proponents of natural selection have been known to use it to "explain" observed biological phenomena in a somewhat after-the-fact manner. Thus, one might say that an animal "does what it does because that particular behavior increases its fitness." Those who succumb to naively explaining away everything as a result of selection have been aptly labeled "adaptationists" (Gould and Lewontin 1979). Used in this way, natural selection can be misleading; it is so pervasive and powerful that nearly any observable phenomenon can be interpreted as a result of selection, even though some must not be. There is a real danger of circularity in such arguments. One should always consider alternative explanations for biological phenomena. Historical factors set various sorts of design constraints on organisms that limit directions in which they can evolve. Perfect adaptation is also prevented by continually changing counterevolutionary pressures from other species. Conflicting demands on allocation of an organism's resources may also preclude ideal solutions to environmental exigencies. These kinds of subtleties will be considered later.
Levels of Approach in Biology
Why do migratory birds fly south in the autumn? A physiologist might tell us that decreasing day length (photoperiod) stimulates hormonal changes that in turn alter bird behavior with an increase in restlessness. Eventually this "Wanderlust" gets the upper hand and the birds head south. In contrast, an evolutionist would most likely explain that, by virtue of reduced winter mortality, those birds that flew south lived longer and therefore left more offspring than their nonmigratory relatives. Over a long period of time, natural selection resulted in intricate patterns of migratory behavior, including the evolution of celestial navigation, by means of differential reproductive success.
The physiologist's answer concerns the mechanism by which avian migratory behavior is influenced by immediate environmental factors, whereas the evolutionist's response is couched in terms of what might be called the strategy by which individual birds have left the most offspring in response to long-term consistent patterns of environmental change (i.e., high winter mortality). The difference between them is in outlook, between thinking in an "ecological" time scale (now time) or in an "evolutionary" time scale (geological time). At the physiologist's level of approach to science the first answer is complete, as is the evolutionist's answer at her or his own level. Mayr (1961) has termed these the "how?" and "why?" approaches to biology. They have also been called the "functional" and "evolutionary" explanations and the " proximate" and " ultimate" factors influencing an event (Baker 1938). Neither is more correct; a really thorough answer to any question must include both, although often only the first can be examined by direct experiment. Nor are those two ways of looking at biological phenomena mutually exclusive; ecological events can always be profitably considered within an evolutionary framework and vice versa.
The evolutionary approach to biological questions is relatively new and has resulted in a major revolution in biology during the last half century. Before then, most biologists merely accepted as immutable a broad range of phenomena, such as the fact that sex ratios are often near equality (50:50), without considering why such facts might be so or how they could have evolved. Although we may not fully understand the causes and consequences of many populational phenomena, we can be confident that most have some sort of evolutionary explanation. This is true of a broad spectrum of observations and facts, such as: (1) some genes are dominant, others are recessive; (2) some organisms live longer than others; (3) some organisms produce many more offspring than others; (4) some organisms are common, others are rare; (5) some organisms are generalists, others are specialists; (6) some species are promiscuous, some polygamous, and some monogamous; (7) some species migrate, others do not; and (8) more species coexist in some areas than in others. All these variables are subject to natural selection. Population biologists are now thinking in an evolutionary time scale, and we have made substantial progress toward a theoretical understanding of why many of the preceding differences occur. Fisher (1930) was one of the first to recognize the power of rigorous application of the genetic theory of natural selection to population biology (sometimes called "selection thinking"), and his book has become a classic. Numerous other biologists have expanded, experimented with, and built on Fisher's groundwork. For example, Lack (1954, 1966, 1968) showed that reproductive rates are subject to natural selection. Others have worked on the evolution of a variety of phenomena, including genetic dominance, mating systems, sex ratio, parental investment, old age, life history patterns, reproductive tactics, foraging tactics, predator-prey interactions, community structure, and so on. Many of these topics, the subject matter of evolutionary ecology, are taken up in subsequent chapters.
Debates and Progress in Ecology
Ecologists are a contentious lot, easily polarized into opposing factions. Early on, there was a separation into the Shelford-Clements community school versus the Gleason individualistic school. During the 1950s and 1960s, Andrewartha and Birch argued for the importance of density independent processes in population regulation, whereas Lack and others emphasized the overriding importance of density dependence. In the 1960s to 1970s, when MacArthur held central stage, resource partitioning studies blossomed. Group selection fell out of vogue in the 1960s, but returned to legitimacy during the 1970s. During the 1970s and 1980s, an acrimonious debate raged over whether or not competition is important in organizing ecological communities. Anyone who invoked competition was labeled dogmatic. The niche concept was seen by some to be useless, or worse. A "predation" school developed which boldly asserted that predation was more important than competition. Simberloff and Strong and their colleagues at Florida State University ("the Tallahassee turks") argued for the "logical primacy" of null models and urged ecologists to adopt a proper Popperian approach to falsifying testable hypotheses. Experimental ecologists express disdain for descriptive studies such as comparative ecology. Statistically inclined ecologists abhor poorly designed experiments, lack of adequate controls, and pseudoreplication (Chapter 14). Some argue about whether or not nature ever reaches an equilibrium, and non-equilibrium perspectives are popular in certain circles. Theoretical ecologists feel that empiricists do not pay enough attention to their precious theories, whereas empirical ecologists do not find dense mathematical abstract treatments of very much interest. Many population ecologists find community ecology to be lacking in rigor.
Healthy debate leads to progress when it forces ecologists to think more rigorously, but deadlock impedes progress. Jargon has sometimes become an end in itself. Vogue has an invisible sway and the pendulum swings back and forth as each generation reacts to the last.
Selected References
Scaling in Biology
Anderson (1986); Brooks (1988); Brown and Maurer (1987, 1989); Holling (1992); Osmund et al. (1980); Pimm (1991); Ricklefs (1987, 1989); Wiens (1989).
Scientific Methods and Human Knowledge
Bender et al. (1984); Colwell and Fuentes (1975); Diamond and Case (1986); Fisher (1935); Hilborn and Stearns (1982); Horn (1979); Hutchinson (1949);
Lakotos (1970); Levins (1966); Loehle (1987); Platt (1964); Popper (1959);
Quinn and Dunham (1983); Schoener (1983).
Domain of Ecology: Definitions and Groundwork
Allee (1951); Allee et al. (1949); Andrewartha (1961); Andrewartha and Birch (1954); Billings (1964); Clarke (1954); Collier et al. (1973); Elton (1927, 1958); Greig-Smith (1964); Harper (1967); Hazen (1964, 1970); Kendeigh (1961); Kershaw (1964); Knight (1965); Krebs (1972); MacArthur (1972); MacArthur and Connell (1966); Macfadyen (1963); May (1973, 1976a); Odum (1959, 1963, 1971); Oosting (1958); Pielou (1969); Platt (1964); Ricklefs (1973); Shelford (1963); Smith (1966); Watt (1973); Whittaker (1970); Wilson and Bossert (1971).
The Urgency of Basic Ecological Research
Ehrlich and Ehrlich (1981); Elton (1927); Lubchenco et al. (1991); Rolston (1985); Weaver and Clements (1929); Wilson (1985).
Natural Selection
Birch and Ehrlich (1967); Darwin (1859); Dobzhansky (1970); Ehrlich and Holm (1963); Emlen (1973); Fisher (1930, 1958a, b); Ford (1964); Gould and Lewontin (1979); Haldane (1932); Kettlewell (1956, 1958); Lande and Arnold (1983); Lewontin (1974); MacArthur (1962); Maynard Smith (1958); Mayr (1959); Mettler and Gregg (1969); Orians (1962); Pianka (1976b); Salthe (1972); Williams (1966a); Wilson and Bossert (1971); Wright (1931).
Levels of Approach in Biology
Baker (1938); Fisher (1930); Lack (1954, 1966, 1968); MacArthur (1959, 1961); MacArthur and Connell (1966); Mayr (1961); Orians (1962); Pianka (1986b).
Debates and Progress in Ecology
Chitty (1996); Conner and Simberloff (1979, 1983); Diamond and Gilpin (1982); Ehrlich and Birch (1967); Gilpin and Diamond (1982, 1984); Hairston, Smith and Slobodkin (1960); Karieva (1989); Murdoch (1966); Quinn and Dunham (1983); Roughgarden (1983); Simberloff (1983); Slobodkin, Smith and Hairston (1967); Strong et al. (1984); Wynne-Edwards (1962).
|