2| History and Biogeography
Self-Replicating Molecular Assemblages
Life began with the first self-replicating molecular assemblage; moreover, natural selection begins to operate as soon as any complex of molecules starts making replicates of itself. No copying device is perfect, and some variants of the molecular assemblage produced are bound to be better than others in their abilities to survive and replicate themselves under particular environmental conditions. As resources become depleted, competition can occur among various self-replicating units. Furthermore, given enough time, some inferior variants presumably become extinct. Thus, each molecular unit maximizes its own numbers at the expense of other such units. Even as the originally simple self-replicating units become more and more elaborate and eventually attain the complex form of present-day organisms, the same principles of natural selection must remain in effect throughout. Thus, we can make certain statements about life that are entirely independent of the precise mechanism of replication. For example, self-replicating molecular assemblages elsewhere in the cosmos doubtlessly will not obey the laws of Mendelian genetics, yet their basic attributes as living material will not be drastically altered. Natural selection and competition are inevitable outgrowths of heritable reproduction in a finite environment. Hence, natural selection exists independently of life on earth. Many of the principles developed here will persist as long as assemblages of molecules replicate themselves anywhere in the cosmos.
Once a self-replicating entity arises, qualitatively new phenomena exist that are not present in an inanimate world. To reproduce, living organisms (or replicating molecular assemblages) must actively gather other materials and energy; that is, they must have some sort of acquisition techniques. Direct and indirect disputes over resources place those units best able to acquire materials and energy (and most successful at transforming them into offspring) at a selective advantage over other such units that are inferior at these processes. Thus, natural selection has the same effect as an efficiency expert, optimizing the allocation of available resources among conflicting demands imposed by foraging, growth, maintenance, and reproduction.
Obviously, the ultimate end point of these processes would be for the one best organismic unit to take over all matter and energy and to exclude all others. This has not occurred for a variety of reasons, as discussed earlier, but especially because of the great variability of the earth's surface, both in time and space. (Humans are making a good stab at it, however!)
The Geological Past
Climatic changes over geological time, called paleoclimatology, are of considerable ecological interest because organisms have had to evolve along with such changes. A really thorough ecological study must include consideration of the past history of the area under study. The earth has changed in innumerable ways during the geological past: the planet's orbital motion undergoes several complex celestial cycles measured on time scales of many thousands of years
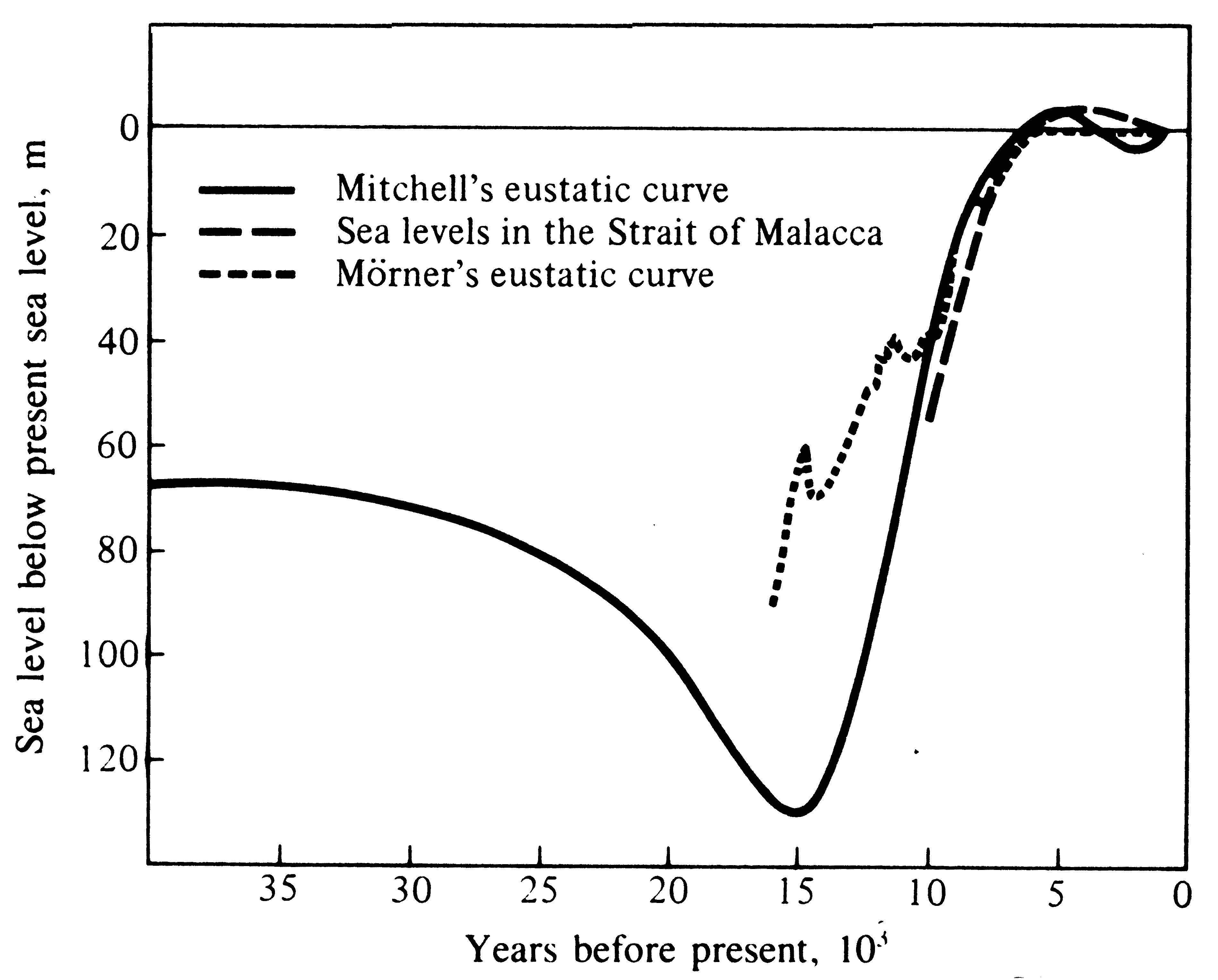
(Milankovitch cycles), its poles have shifted and wandered, periods of orogeny (mountain building) by tectonic upheaval of the earth's crust have waxed and waned at different places and times, the continents have "drifted" and moved on its mantle, and the planet itself has alternately warmed and cooled, the latter resulting in periods of extensive glaciation. Sea levels dropped during glacial periods as water accumulated on land as snow and ice (such sea level changes controlled by glacier alterations are called "eustatic"; see Figure 2.1).
-
Figure 2.1. Estimates of sea level changes over the past 35,000 years from three different sources.
[After Williamson (1981) by permission of Oxford University Press.]
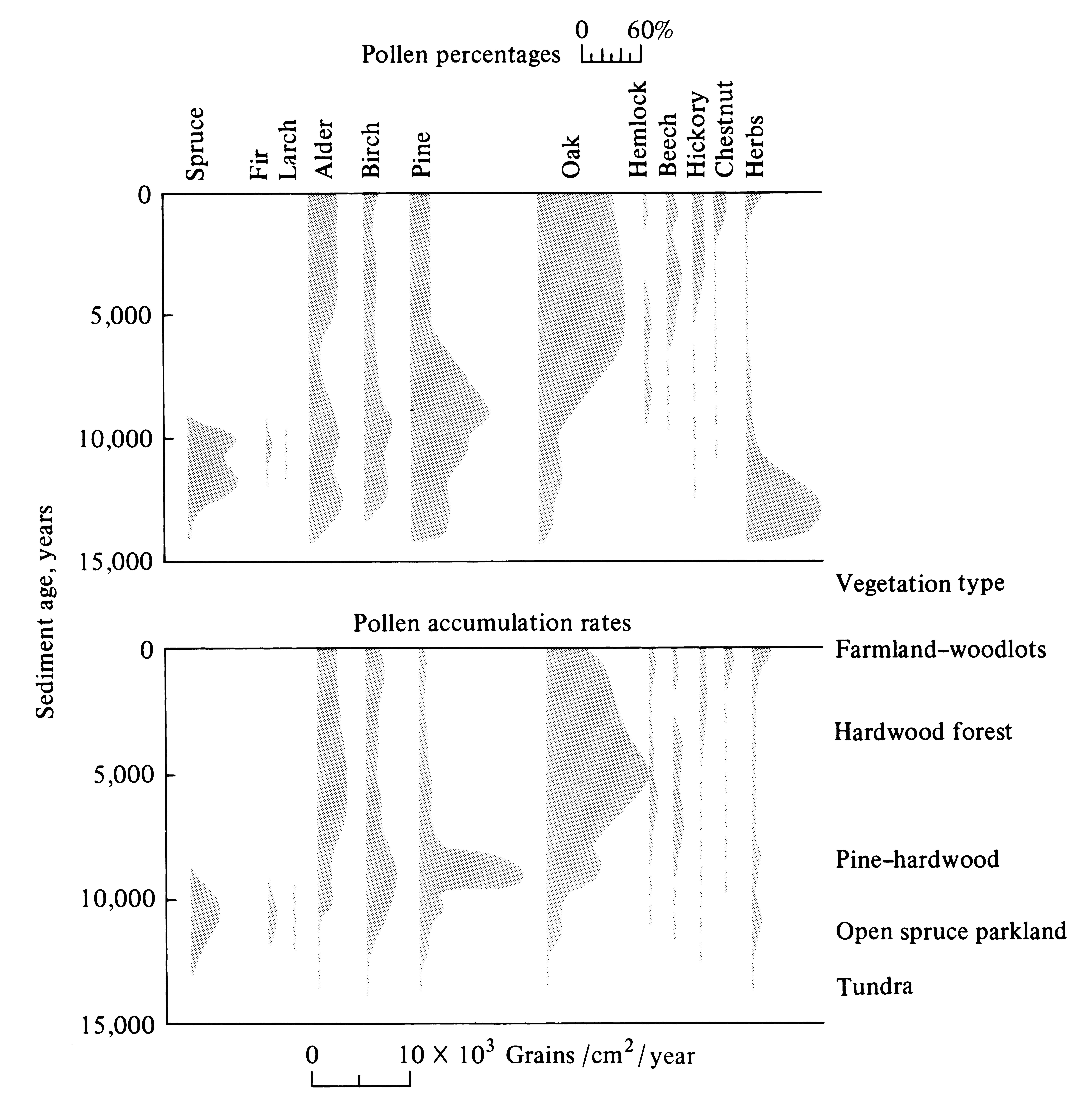
Although it is difficult to trace such distant history, a variety of techniques, some of them quite ingenious, have been developed that allow us to deduce many of the changes that earth has undergone. One way to look into the past is to examine the fossil record. Lake sediments are an ideal source of layered fossils and have often been used to follow the history of an area. Fossilized pollens in a lake's sediments are relatively easily identified; pollens of plants adapted to particular types of climates can be used as indicators of past climates, as well as the types and composition of forests that prevailed near the lake at different times (see Figure 2.2). Of course, such palynological analyses are fraught with difficulties due to both variations between taxa in rates of pollen production and differential transport and deposition. Moreover, many deposits may contain mixtures of pollens from several different communities.
-
Figure 2.2. Fossil pollen profiles from dated layers of lake sediments in northeastern United States for the period following the last ice age. Upper plot shows number of pollen grains of each species group as a percentage of the total sample. Lower plot gives estimated rates of deposition of each type of pollen and, at the right, the type of vegetation that probably prevailed in the area. [Adapted from Odum (1971) after M. Davis.]
A technique known as carbon dating allows estimation of the age of fossil plant remains, including pollens and charcoal. Solar radiation converts some atmospheric nitrogen into a radioactive isotope of carbon called carbon 14 (14C). This 14C is oxidized to carbon dioxide and is taken up by plants in photosynthesis in proportion to its abundance in the air around the plant. All radioactive isotopes emit neutrons and electrons, eventually decaying into nonradioactive isotopes. Half of a quantity of 14C becomes nonradioactive carbon 12 (12C) each 5600 years (this is the half-life of 14C). When a plant dies, it contains a certain maximal amount of 14C. Comparison of the relative amounts of 14C and 12C in modern-day and fossil plants allows estimation of the age of a fossil. Thus, a fossil plant with half the 14C content of a modern plant is about 5600 years old, one with one-quarter as much 14C is 11,200 years old, and so on. The carbon dating method has been checked against ancient Egyptian relics of known age made from plant materials; it accurately estimates their ages, confirming that the rate of production of 14C and the proportion of 14C to 12C have not changed much over the last 5000 years. The technique allows assignment of fairly accurate ages to all sorts of recently fossilized plant materials. Other methods of radiometric dating, such as uranium-lead, uranium-thorium, and potassium-argon dating, allow older materials to be dated.
A similar dating technique makes use of the observation that the uptake of two oxygen isotopes, 16O and 18O, into carbonates is temperature dependent. Thus, the proportion of these two isotopes in a fossil seashell presumably reflects the temperature of an ancient ocean in which that particular mollusk lived.
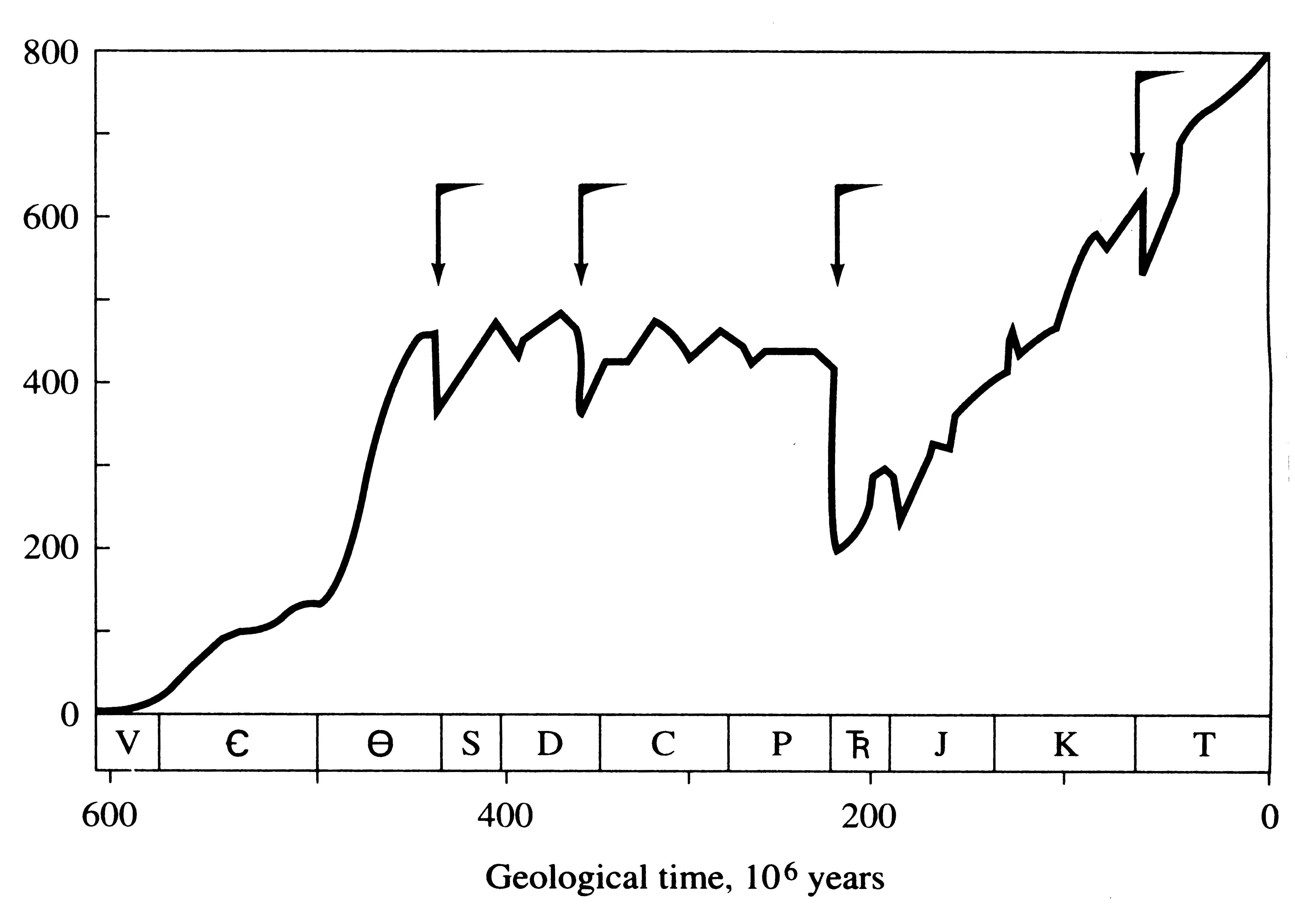
The geological time scale is summarized in Table 2.1. Several major episodes of extinction stand out in the fossil record (Figure 2.3). These events are so striking
that they are used to mark the boundaries between geological periods: (1) 70 percent of marine life went extinct at the Paleozoic-Mesozoic boundary, (2) the last dinosaurs went extinct at the Mesozoic-Tertiary transition, and (3) over
many different parts of the planet, numerous species of large mammals died out rather suddenly and dramatically near the end of the Pleistocene ice ages.
Table 2.1 The Geological Time Scale
______________________________________________________
 Years in millions Years in millions
since beginning of
Eras Periods Epochs period or epoch
__________________________________________
Quaternary Recent0.1
Pleistocene 1.6
___________________________________________
CenozoicPliocene5
Miocene 22
Tertiary Oligocene36
Eocene 55
Paleocene 65
_______________________________________________________
Cretaceous144
MesozoicJurassic192
Triassic245
_______________________________________________________
Permian290
Carboniferous360
Devonian408
PaleozoicSilurian435
Ordivician485
Cambrian570
_______________________________________________________
Precambrian4600
_______________________________________________________
Long geological periods without major extinctions, followed by abrupt periods of massive extinctions, are known as "punctuated equilibria." These widespread extinctions markedly altered almost all ecological communities. Moreover, their synchrony demands attention, since it strongly suggests general underlying causal explanations. Perhaps the most intriguing of those suggested for the extinction of the dinosaurs is the asteroid impact hypothesis (Alvarez et al. 1980), which asserts that extraterrestrial dust from a large comet drastically decreased incident solar radiation and hence primary productivity (this would also have made the planet colder). In support of this hypothesis, a thin fossil layer of iridium, a rare element on the earth but one that is more abundant in meteorites, has been found at widespread localities around the planet dating from this period.
Intriguing speculation has also been offered for the Pleistocene mammal extinctions (Martin and Wright 1967), including the possibility of "overkill" by prehistoric humans (Martin 1967), although climatological events must certainly have played a major role as well (Guilday 1967; Graham 1986). Pleistocene assemblages of small mammals seem to have been more diverse and fundamentally different than modern ones (Graham 1986).
-
Figure 2.3. Marine extinctions in the fossil record. V = Precambrian, C = Cambrian,
O = Ordivician,
S = Silurian, D = Devonian, C = Carboniferous, P = Permian, TR = Triassic,
J = Jurassic, K = Cretaceous, T = Tertiary (see Table 2.1) [After Erwin et al. (1987).]
South America was virtually completely isolated for most of the past 70 million years. In the absence of many major groups of placentals, a rich fauna of marsupials, including a saber-toothed catlike predator, evolved over this long time period. Edentates and proto-ungulates were present, and a rodent, a monkey, and a small raccoon-like carnivore did eventually reach the isolated continent. When the isthmus of Panama was formed about 3 million years ago, numerous different taxa of land and freshwater organisms were exchanged between North America and South America (this process has been termed the "great American biotic interchange"). Although Central America has since come to be dominated by South American elements, the impact of the North American fauna on South America was much more profound than the effect of the South American biota on that of North America. Many South American marsupial species went extinct, although a few ancient lineages persisted. South American mammals that have successfully invaded North America include the armadillo, opossum, and porcupine.
Reasons for many of the earth's past changes, such as polar movements and the alternate warming and cooling of the planet, are little known and may well involve solar changes. Piecing together all this diverse and sometimes conflicting evidence on the earth's past history is a most difficult and challenging task and one that occupies many fine minds.
Classical Biogeography
A major goal of ecology is to understand various factors influencing the present distribution and abundance of animals and plants (Andrewartha and Birch 1954; Krebs 1972; MacArthur 1972). Factors affecting abundances and microgeographic distributions (including habitat selection) will be considered later; here, we examine more gross geographic distributions -- the spatial distributions of plants and animals over large geographic areas such as major landmasses (continents and islands). The study of the geographical distributions of plants and animals, respectively, are termed phytogeography and zoogeography. Biogeography encompasses the geography of all organisms and involves a search for patterns in the distributions of plants and animals and an attempt to explain how such patterns arose during the geological past. In addition to classifying present distributions, biogeographers seek to interpret and to understand past movements of organisms. Ecology and biogeography are closely related and overlapping disciplines and have profoundly affected one another.
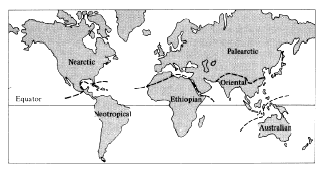
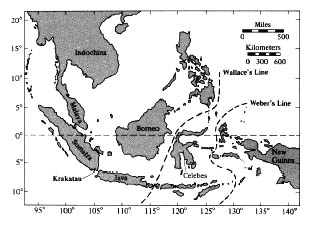
When early naturalists traveled to different parts of the world, they quickly discovered distinctly different assemblages of species. As data were gathered on these patterns, six major biogeographic "realms" or regions were recognized, three of which correspond roughly to the continents of Australia (Australian), North America north of the Mexican escarpment (Nearctic), and South America south of the Mexican escarpment (Neotropical). (The Neotropical region also includes the Antilles.) Africa south of the Sahara is known as the Ethiopian region. Eurasia is divided into two regions, the Palearctic north of the Himalayas (which includes Africa north of the Sahara desert) and the Oriental south of the Himalayas (India, southern China, Indochina, the Philippines, and Borneo, Java, Sumatra, and other islands of Indonesia east to, and including, the Celebes). Each of the six biogeographic regions (Figure 2.4) is separated from the others by a major barrier to the dispersal of plants and animals, such as a narrow isthmus, high mountains, a desert, an ocean, or an oceanic strait. There is generally a high degree of floral and faunal consistency within regions and a marked shift in higher taxa such as genera and families in going from one region to another. Although biogeographers familiar with different plant and animal groups often disagree on the exact boundaries between regions (Figure 2.5), there is broad agreement on the usefulness of recognizing these six major regions. High species diversities in the tropics, among other things, have led to the notion that speciation rates in these areas must be extremely high and that such regions often constitute "source areas" for production of new species, many of which then migrate into less hospitable areas, such as the temperate zones. Thus, Darlington (1957, 1959) proposed the "area-climate hypothesis," which states that most dominant animal species have arisen in geographically extensive and climatically
-
Figure 2.4. The six major biogeographic regions of the world.
-
Figure 2.5. Wallace's and Weber's "lines" in southeast Asia, which separate the Oriental from the Australian regions. The position of the volcanic island of Krakatau between Sumatra and Java is indicated at the lower left.
favorable areas; he considered the Old World Tropics, which include the tropical portions of the Ethiopian and Oriental regions, to be the major source area for most vertebrate groups and argued that such dominant forms have migrated centrifugally to other smaller and less favorable areas, including Europe, North and South America, and Australia.
Classical biogeography has produced several so-called biogeographic rules based on recurring patterns of adaptation of organisms. Thus, homeotherms living in cold climates are often larger than those from warmer regions. Such a trend or cline can even be demonstrated within some wide-ranging species. This tendency, termed Bergmann's rule after its discoverer, has a probable causal basis in that large animals have less surface per unit of body volume than small ones, resulting in more efficient retention of body heat. Many other biogeographic rules have also been proposed, all of which are basically descriptive. Allen's rule states that the appendages and/or extremities of homeotherms are either longer or have a larger area in warmer climates; a jackrabbit, for example, has much longer and larger ears than an arctic hare. The presumed functional significance is that large appendages, having a larger relative surface area, are better at heat dissipation than smaller ones. Another rule (Gloger's) asserts that animals from hot, dry areas tend to be paler than those from colder, wetter regions. Still another biogeographic rule is that fish from colder waters often have more vertebrae than those from warmer waters. The adaptive significance of many of these biogeographic trends remains obscure, although such geographically variable phenotypic traits are frequently developmentally flexible and respond more or less directly to temperature.
Continental Drift
Much of this classical biogeography assumed some permanence in the locations of continents. As a result, interpretations of faunal similarities between them often relied on hypothetical mechanisms of transport from one continent to another, such as "rafting" of organisms across water gaps. Such long-distance dispersal events are exceedingly improbable, although they must occur from time to time (cattle egrets, for example, apparently made a successful transatlantic crossing from Africa to South America without human assistance late in the 18th century).
Massive movements of the continents themselves were first suggested by Taylor (1910), but his bold hypothesis was not widely accepted. Wegener is usually credited with
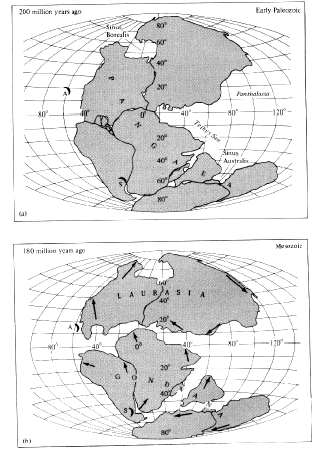
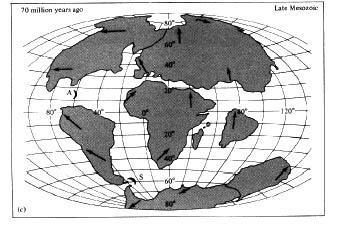
the idea of continental drift. Recent advances in geology indicate that the continents were once joined in a large southern landmass (Pangaea) and have "drifted" apart (Figure 2.6), with a gradual breakup that began in the early Mesozoic era (about 200 million years ago). Geological evidence that the continents have drifted (and are now drifting) is now extensive (Hallam 1973; Marvin 1973; J. T. Wilson 1973; Skinner 1986). Smaller fragments of land, known as "suspect terraines," also appear to have moved.
-
Figure 2.6. Approximate positions of major landmasses at different times in the geological past, showing their probable movements. [Adapted from Dietz and Holden (1970). The Breakup of Pangaea. Copyright © 1970 by Scientific American, Inc. All rights reserved.]
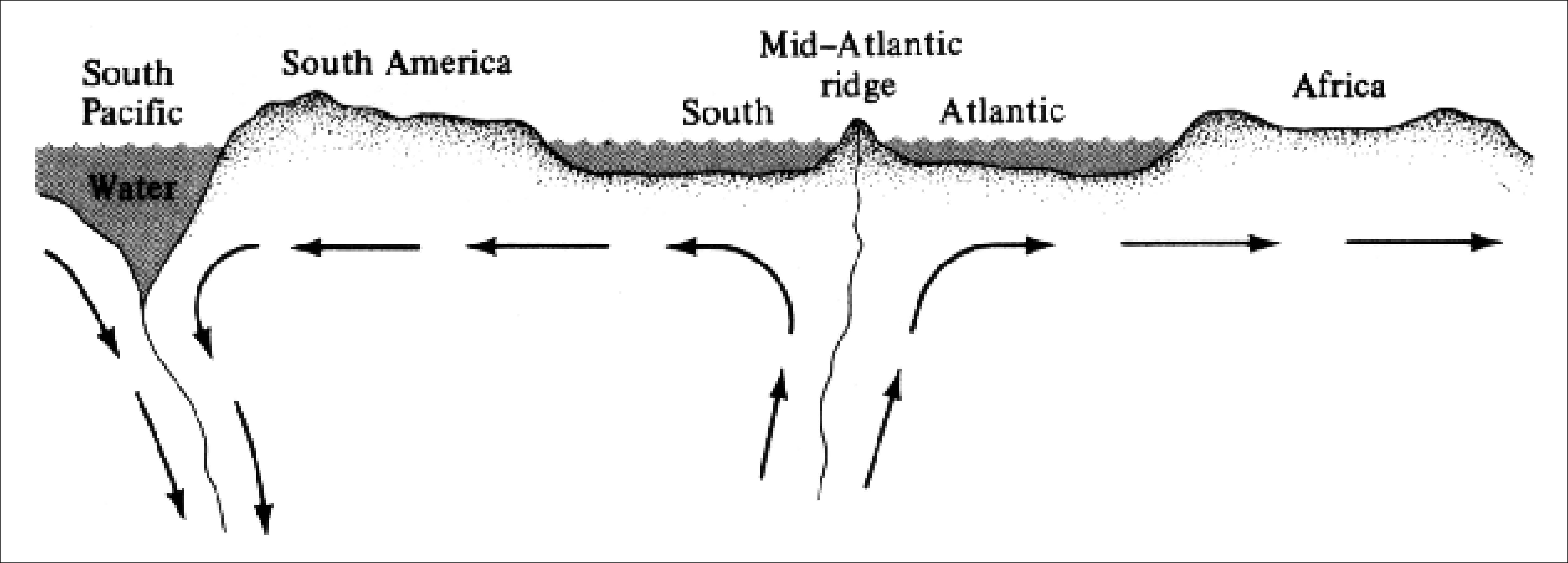
Certain types of rocks, particularly basalts, retain a magnetic "memory" of the latitude in which they were solidified. Such paleomagnetic evidence allows mapping of the past position of the North Pole. (An intriguing but as yet still unexplained phenomenon has been discovered: reversals in magnetic polarity occur in geological time, the last of which has been dated at 700,000 years ago.) Recent rocks from different locations coincide in pinpointing the pole's position, but paleomagnetic records from older rocks from different localities are in discord. These discrepancies strongly suggest that the continents have moved with respect to one another. The continents are formed of light "plates" of siliceous, largely granitic, rocks about 30 km thick, which in turn float on denser mantlelike basaltic blocks. The ocean floors are composed of a relatively thin altered top of the earth's mantle. A mountain range of the seafloor in the mid-Atlantic represents a region of upwelling of the mantle. Under this interpretation, as the upwelling proceeds, seafloors spread and continents move apart (Figure 2.7). The positions of paleomagnetic anomalies (polarity reversals) in the seafloor allow geologists to calculate the velocity of lateral motion of the ocean floors, which correspond to comparable estimates for the landmasses. Thus, modern theory holds that, except for the Pacific (which is shrinking), the oceans are growing, with very young ocean floors in midocean and progressively older floors toward the continents. Other evidence, such as the apparent ages of islands and the depths of sediments, nicely corroborate this conclusion.
-
Figure 2.7. Diagrammatic cross-sectional view of the probable movements of the earth's mantle and crust that led to seafloor spreading and continental drift. Upwelling of deep mantle materials in the mid-Atlantic is accompanied by a surface movement away from the mid-oceanic ridge. The continents, which float on top of these moving denser materials, are carried along. At the oceanic trench on the far left, these materials sink back down a subducting zone into the mantle, forming a closed system of circulating materials.
Much of classical biogeography is being reinterpreted in light of these new findings. For example, certain very ancient groups of plants, freshwater lungfishes, amphibians, and insects that had spread before the breakup of the continents now occur on several continents, whereas many other more recently evolved groups of plants and animals, such as mammals and birds, are restricted to particular biogeographic regions. These latter, more recent groups follow the regional divisions much more closely than the older groups (Kurtén 1969).
Imagine the effects of changing climates on plants and animals as continents drift through different latitudes! The Indian subcontinent changed from south temperate to tropical to north temperate and was virtually a "Noah's ark" carrying a flora and fauna. Australia became arid as it drifted into the mid latitudes; moreover, as this landmass continues to move toward the equator, its climate will gradually become wetter and more tropical.
Selected References
Self-Replicating Molecular Assemblages
Bernal (1967); Blum (1968); Calvin (1969); Ehrlich and Holm (1963); Fox and Dose (1972); Oparin (1957); Ponnamperuma (1972); Salthe (1972); Wald (1964).
The Geological Past
Alvarez et al. (1980); Birch and Ehrlich (1967); Brown (1982); Dansereau (1957); Darlington (1957, 1965); Dietz and Holden (1970); Erwin et al. (1987); Graham (1986); Guilday (1967); Hesse, Allee, and Schmidt (1951); Imbrie et al. (1984); Jelgersma (1966); Martin (1967); Martin and Klein (1984); Martin and Mehringer (1965); Martin and Wright (1967); Olsen (1983); Sawyer (1966); Skinner (1986); Stehli and Webb (1985); Tarling (1983); Udvardy (1969); Wilson (1971, 1973); Wiseman (1966); Wright and Frey (1965).
Classical Biogeography
Andrewartha and Birch (1954); Cain (1944); Dansereau (1957); Darlington (1957, 1959, 1965); Grinnell (1924); Hesse, Allee, and Schmidt (1951); Krebs (1972); MacArthur (1959, 1972); MacArthur and Wilson (1967); Nelson and Platnick (1981); Newbigin (1936); Schall and Pianka (1978); Terborgh (1971); Udvardy (1969); Wallace (1876); Watts (1971).
Continental Drift
Cracraft (1974); Dietz and Holden (1970); DuToit (1937); Kurtén (1969); Hallam (1973); Marvin (1973); Skinner (1986); Tarling and Runcorn (1975); Skinner (1986); Taylor (1910); Wegener (1924); J. T. Wilson (1971, 1973).
|