|
Pianka has exploited lizards as exemplars and model organisms to study a wide variety of basic ecological phenomena. In addition to becoming the world's leading authority on the ecology of desert lizards, he has become one of its best-known evolutionary ecologists. His contributions can be broken down into five major categories, as follows: 1. Natural History and Ecology of Lizards  a. North American Flatland Desert Lizards
b. Australian Desert Lizards
c. Kalahari Desert Lizards
d. Intercontinental Comparisons
e. Periodic Table of Lizard Niches a. North American Flatland Desert Lizards
b. Australian Desert Lizards
c. Kalahari Desert Lizards
d. Intercontinental Comparisons
e. Periodic Table of Lizard Niches
2. Evolutionary Ecology a. Foraging Strategies
b. Reproductive Tactics
c. Competition and Niche Theory
d. Community Structure and Organization
e. Species Diversity
3. Landscape Ecology a. Fire Succession
b. Spatial-Temporal Habitat Mosaics
c. Habitat Fragmentation-Diversity Paradox
d. Metapopulation Structure
4. Biogeography a. Geographical Patterns in Diversity
b. Zoogeography of Australian Desert Lizards
c. Theoretical Biogeography
5. Phylogenetics and the Modern Comparative Method a. Evolution of Body Size in Varanid Lizards
b. Adaptive Radiation of Australian Skinks
As a graduate student at the University of Washington during 1960-65, Pianka identified lizards as nearly ideal animals for ecological research. Because lizards are ectotherms, they are ëlowí energy animals. Lizards are often abundant, making them relatively easy to locate, observe, and capture. By facilitating metabolic inactivity on both a daily and a seasonal basis, ectothermy may well confer lizards with an advantage over birds under conditions of low and unpredictable productivity such as in desert regions. Many lizard species are low in mobility and hardy in captivity. In addition, izards eat their prey as whole intact items, which greatly facilitates analysis of diets via stomach contents (birds and mammals break or grind their food up into small pieces). Lizards exhibit a wide range of variation in many morphological, behavioral, physiological, and ecological phenomena. Some species of lizards are annuals, whereas others live for decades. Reproductive tactics are readily studied and quite varied. Some species, such as geckos and the iguanian Anolis, have invariate s mall clutch sizes. Other species are exceedingly prolific, laying large clutches of several dozen eggs. Viviparity has arisen repeatedly among different lineages, as have fringed toe lamellae. Both arboreal and terrestrial lizards occur among both nocturnal and diurnal species. Some species are highly aquatic as well. Lizards inhabit a broad range of habitats, including deserts, grasslands, chaparral, rock outcrops, deciduous forest and rainforest. Lizards exhibit a wide range of thermoregulatory tactics, ranging along a continuum from passive thermoconformers to active thermoregulators. Until the 1960's, birds had dominated ecology as study organisms, but work of Pianka and others has changed that. Pianka's doctoral dissertation research was on the natural history and ecology of North American desert lizards, with particular reference to factors influencing species diversity. This detailed 3-year field study of a series of ten flatland desert sites ranging from southern Idaho across nearly a 1,000 mile latitudinal transect to southern Arizona demonstrated that the structural complexity of the vegetation was the primary factor influencing the numbers of species of lizards present. Pianka's doctoral committee (Drs. R. C. Snyder, R. T. Paine, and G. H. Orians) clearly recognized the importance of his early contributions, summarizing his Ph. D. research effort in 1965 as follows:
In addition to two important publications on the problem of factors determining diversity of lizard species in the flatland deserts of western North America (Pianka 1966, 1967), he exploited his data in greater detail to examine the comparative autecology of the teid Cnemidophorus (now Aspidoscelis) tigris in different parts of its geographic range (Pianka 1970). He found that a whole host of ecological phenomena varied with latitude, including diets, activity patterns, thermoregulation and predation. In the northern parts of this lizard species' geographic range, physical factors are of overriding importance, whereas in the south, biotic factors, particularly predation, assume paramount importance. In collaboration with W. S. Parker, he published a series of half a dozen papers on the natural history and ecology of various other lizard species (Pianka and Parker 1972, 1975; Parker and Pianka 1973, 1974, 1975, 1976). One of Pianka's ex-graduate students, Jos. J. Schall (now a full Professor at the University of Vermont), studied several species of Cnemidophorus (now Aspidoscelis) in west Texas, including some unisexual all-female species. Finding their data to be complementary, they co-authored a paper on the diversity of escape tactics in whiptail lizards (Schall and Pianka 1980). While studying diversity and ecology of North American desert lizards, Pianka conceived an ideal follow-up research project: a natural experiment to compare an independently evolved desert-lizard system with North America. Australian desert lizards were virtually unknown at the time. He was awarded a 3-year NIH postdoctoral fellowship to work with the late R. H. MacArthur at Princeton University where he applied for a supplementary grant from NSF to undertake studies on Australian desert lizards. Pianka pioneered difficult field work in the outback and rediscovered what aborgines knew, making it scientific and quantitative. Pianka spent 18 months in Australia under to sponsorship of A. R. Main (Zoology, University of Western Australia), where he discovered and popularized Earth's richest known lizard assemblages. In the Great Victoria desert of Western Australia, as many as 55 species of lizards occur in sympatry on sandridge study sites, more than twice as many species as are found together in similar habitats in other parts of the world. The challenge of understanding these incredibly diverse saurofaunas was awesome and daunting. In 1966-68, Pianka and his ex-wife Helen discovered seven new species of Ctenotus skinks, which were described by Dr. Glen Storr of the W. A. Museum 40 years ago. Storr honored them by naming one species after her (helenae) and another after him (piankai). Pianka published a detailed study of the comparative autecology of 14 species of desert Ctenotus in 1969 (Pianka 1969). He also published extensive natural history data, including information on diet, thermoregulation, and reproductive tactics on dozens of species of agamids, geckos, pygopodids, skinks, and varanids. His specimen collections are deposited in major museums. Pianka discovered and described a rather substantial degree of habitat specificity among Australian desert lizards, and he interpreted the significance of this phenomenon in general terms. He discovered and published many major range extensions. Indeed, he described the vertebrate fauna of the Great Victoria desert for the first time. Pianka proposed a synthetic scheme for understanding the biogeography and speciation of Australian desert lizards, which included identifying several major routes by which habitat-specialized lizards could disperse between regions. He named an important east-west strip of shrub-mulga habitat across the center the Giles Corridor (Pianka 1972). Indeed, his zoogeography paper is still considered state-of-the-art a quarter of a century later! Pianka has since returned to the Great Victoria Desert to continue his long-term studies many times, as a Guggenheim Fellow (1978-79), as a Fulbright Senior Research Scholar (1990-91), and under Faculty Research Assignments from the University of Texas at Austin (1989-90, 1992, 1995-96, 1998, 2003, 2008). Pianka (1996) published a chapter entitled Long-term changes in Lizard Assemblages in the Great Victoria Desert. Pianka has lived five full years of his life in the Australian deserts and is recognized as the world's authority on their lizard fauna. The great disparity between lizard diversity in North America (4-11 species) and Australia (18-55 species) made meaningful comparisons difficult. The Kalahari desert-lizard system is intermediate in diversity with 12-17 species. Pianka obtained an NSF grant to study the ecology of Kalahari lizards in 1969-70 and employed Raymond B. Huey (now a Professor at the University of Washington, Seattle) as a field research assistant to spend a full year acquiring a data set for comparison with those from North America and Australia. Among the highlights of the Kalahari research was the discovery and documentation of character displacement in termite-specialized fossorial Typhlosaurus skinks (Huey and Pianka 1974; Huey, Pianka, Egan, and Coons, 1974). Huey and Pianka (1977) also described a remarkable case of Batesian mimicry in which a juvenile lizard mimics a noxious beetle. Another paper examined evidence for thermoregulation among diurnal Kalahari lizards (Huey and Pianka 1977). Still another analyzed resource partitioning among nocturnal Kalahari geckos (Pianka and Huey 1978). Pianka expected to find patterns in Australian deserts similar to those he had seen in North America. He was especially interested in making detailed comparisons of so-called ecological equivalents, independently evolved lineages that occupy roughly the same ecological roles on different continents. A prime example is North American horned lizards Phrynosoma, which are in a different family than their Australian counterpart the thorny devil Moloch horridus. Both are cryptically-colored and thornily-armored ant-specialized species: an iguanian Phrynosoma platyrhinos exploits this ecological role in North America, while its counterpart the agamid Moloch occupies it in Australia. No Kalahari lizard has adopted such a life style. 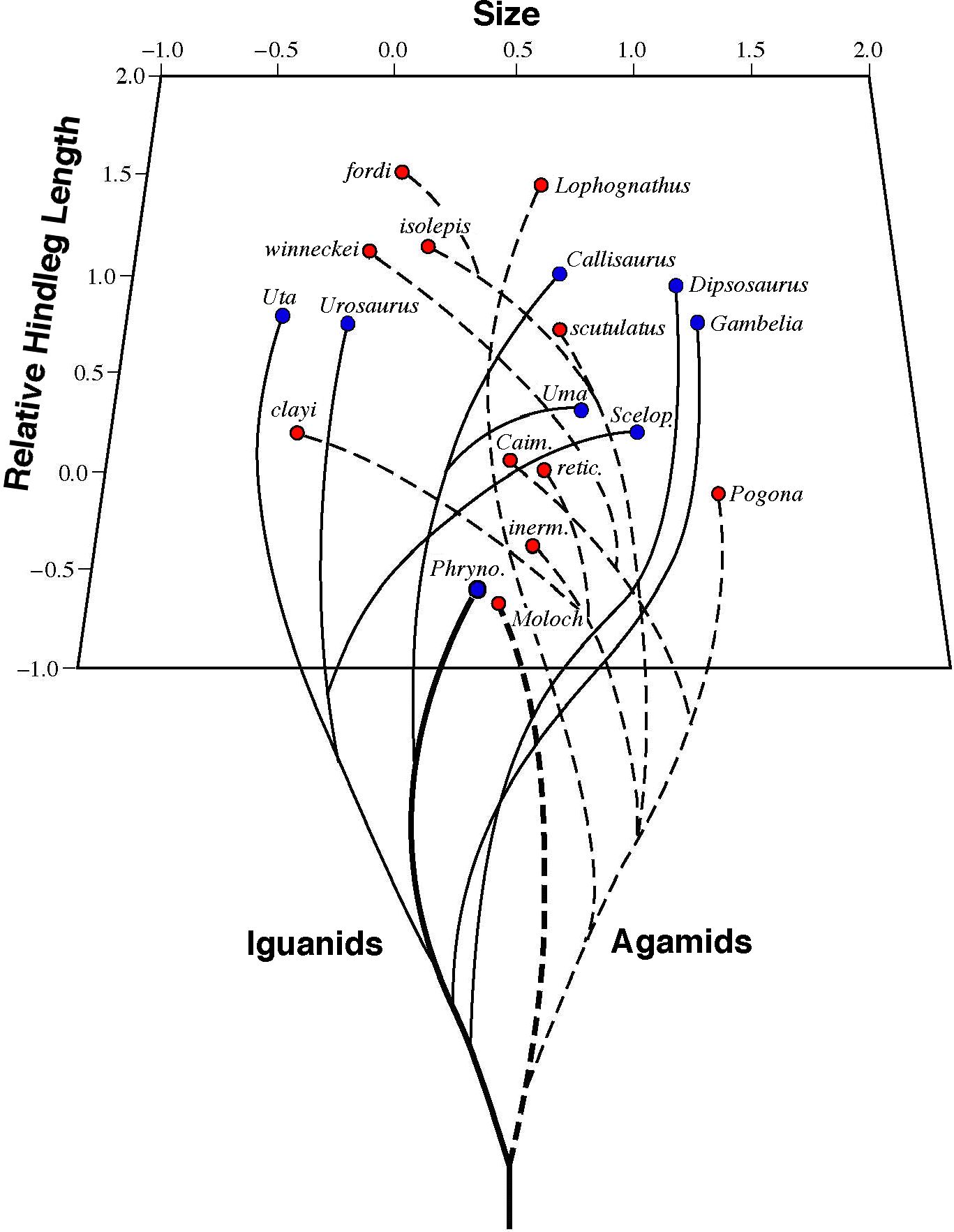
Pianka pioneered intercontinental comparative ecology, from the microecological to faunal levels, and provided ecology with a textbook panglobal example of the importance of historical processes. He analyzed the structure and organization of lizard communities and demonstrated that differences in niche breadth are not a primary factor contributing to differences in diversity, but that major factors are niche dimensionality and the ranges of resources utilized. His 1973 review of the structure of lizard communities was chosen as a Citation Classic and his 1974 paper on niche overlap and diffuse competition was reprinted in Niche: Theory and Application. Benchmark Papers in Ecology. Intercontinental comparisons of desert-lizard assemblages from three independently-evolved systems with differing diversities demonstrate no differences in niche breadth, which is often bimodally distributed among species suggesting a natural dichotomy of generalists and specialists (Pianka 1973, 1985, 1995). In 1986, Pianka published a synthesis of his life's work on the comparative ecology of desert lizards in three comparable but independently-evolved ecological systems. Study of such widely-spaced systems by the same researcher using identical field techniques provides unique material for understanding questions of community organization and ecological convergence. Among other analyses, he generated many interesting pseudo-communities which were compared against their real prototypes to detect structure. He also used these data to perform a massive series of computer-generated experiments involving thousands of reciprocal species transplants among study sites. Aliens did not mesh as well with other species in a given system as did residents, suggesting that these natural ecological systems are organized. Eric R. Pianka, Laurie J. Vitt, Nicolas Pelegrin, Daniel B. Fitzgerald, and Kirk O. Winemiller Ecologists begin to construct a Periodic Table of Niches Download pdf 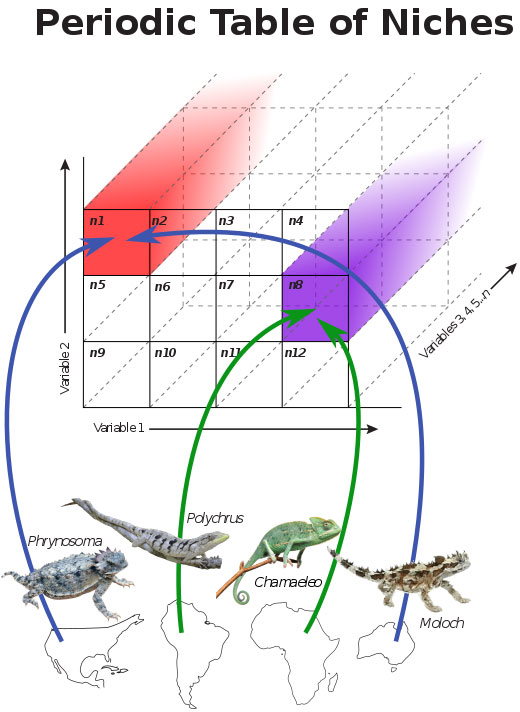 Five prominent ecologists present new insights into a concept first suggested by the late Robert H. MacArthur over half a century ago. MacArthur suggested a Periodic Table of ecological niches could be constructed similar to the chemist's Periodic Table of Elements, which are ordered by a combination of their atomic number (protons), configuration of electrons, and certain chemical properties. Throughout, the paper features a back-and-forth interplay among outright natural history, niche theory, state-of-the-art multivariate statistical analyses, and downloadable rotatable 3D plots. Ecological niches are so complex that the notion they could somehow be ordered in a predictable way has eluded ecologists. The niche defines how species live, reproduce, and interact with other organisms and their environments. Consequently, niches are multidimensional. Visualizing and understanding such complex systems is difficult because humans perceive only three dimensions. In a major synthesis appearing in The American Naturalist, a niche ordination and classification framework is presented based on extensive ecological data gathered by the first two authors during the course of two lifetimes of fieldwork conducted in diverse habitats on four continents over the last half-century. A key prediction of this theory is that distantly related species on different continents should evolve to fill identifiably similar niches, and should exhibit similar sets of ecological traits, referred to as evolutionarily convergence by ecologists. To test this prediction, data were assembled on five major niche dimensions (habitat, diet, life history, metabolism, and defense, each with 7-15 variables) for 134 species representing 24 of the 38 extant lizard families. Lizard niche dimensions have evolved in concert; transitions in life history and trophic traits occur in synchrony. Natural dichotomies include activity time, foraging mode, parity mode (egg laying/live bearing), and habitat. Patterns are repeated: Australian desert lizards solve problems in the same ways that African and American lizards do, even when they are not evolutionarily related.  Convergent evolution among lizards around the world is one of the pillars for the idea of a Periodic Table. Based on multivariate analyses of 50+ lizard niche dimensions, just three capture 61.7 % of the total variation indicating that lizard niches are tightly constrained. To more easily visualize their results, the authors produced rotating 3-dimensional graphics (link to 3-dimensional graphics) that allow exploration of constraints and tradeoffs in the evolution of lizard niches. These reveal the manner in which species overlap or separate out based on habitat, body size, foraging mode, diet, life history, metabolism, defensive tactics, and/or time and place of activity. One hundred distantly-related convergent species pairs from different geographic regions stay close to each other as niche space is rotated ( link to 3D plots showing 50+ dimensions, lizard niche hypervolume, some convergent pairs, and various other dimensions), thus confirming periodicity in lizard niche dimensions, and strongly supporting the utility of a Periodic Table. Extending this approach to other taxa should lead to a wider understanding of niche evolution. Events and phenomena of interest in ecology can be interpreted and understood at two levels: the functional and evolutionary approaches, which neatly complement each other. Both levels of approach are required for a really thorough understanding and appreciation of any particular ecological phenomenon. Rigorous application of the theory of natural selection has provided powerful new insights into the evolution of a wide range of ecological phenomena, including sex ratios, mating systems, social behavior, senescence, patterns and modes of foraging, reproductive tactics and life histories, competitive interactions, niche relationships, predator-prey interactions (including aspect diversity and plant-herbivore interactions), coevolution, community structure, indirect interactions among species in complex networks, metapopulation structure, and ecological succession. As a graduate student, Pianka wrote two important papers which were published in The American Naturalist. One, co-authored with fellow graduate student Mary F. Willson, was a short but thoughtful essay on the interrelationships among sexual selection, sex ratio, and mating systems. The other was the first comprehensive major review of factors affecting latitudinal gradients in species diversity (now known as biodiversity). This review, published in 1966, was reprinted in 1981 in Tropical Ecology. Benchmark Papers n Ecology. Pianka's conceptual contributions are wide ranging and include foraging theory (he coined the terms sit-and-wait versus widely foraging and he coauthored the first paper on optimal foraging), reproductive tactics (he popularized r and K selection and made the esoteric notion of residual reproductive value easily understandable to the lay person), allocation theory and optimality (he embraced the notion of trade-offs from the very beginning), intercontinental comparisons (he pioneered such studies), resource partitioning (he developed the protocols), community structure (he wrote several important reviews on this subject), species diversity (his forte), and, more recently, biogeography, landscape ecology, metapopulation structure, and phylogenetic systematics. Pianka invented many new techniques and concepts -- his publications include four Citation Classics, which have changed the way most ecologists think forever. Many of his papers have been reprinted in edited collections of classic papers in ecology. Pianka has maintained a keen interest in general questions in evolutionary ecology and his teaching efforts have been directed towards this area. He has written numerous papers and chapters of general interest, including his well-known textbook Evolutionary Ecology (another Citation Classic), which underscores his leadership in ecology. This book persisted over three decades (through 6 editionsand has been translated into Greek, Japanese, Polish, Russian, and Spanish and is now available as an eBook. An entire generation of ecologists have been educated from this very important book. Pianka has also published several other highly significant books, including two coedited symposium volumes on lizard ecology, a synthesis of his life's research, plus a recent intriguing autobiographical account of the adventures facing a desert naturalist (The Lizard Man Speaks, University of Texas Press). With coauthor Laurie Vitt, he wrote the most important book ever written on lizards Lizards: Windows to the Evolution of Diversity published by the University of California Press in 2003. This book won Best non-fiction book, Oklahoma Center for the Book in 2004 and the Grand Prize at the Ninth Annual UT Coop Robert W. Hamilton Book Awards in 2005. 2a. Foraging Strategies Earnest enquiries into animal foraging behaviors began three decades ago with publication of three important papers (MacArthur and Pianka 1966, Emlen 1966, and Pianka 1966). The first two papers by MacArthur and Pianka (1966) and Emlen (1966) were theoretical, examining the logic of animal feeding behaviours by identifying benefits and costs associated with various activities. These seminal papers established the field now known as optimal foraging theory (OFT). MacArthur and Pianka (1966) is also a Citation Classic and was reprinted in 1991 in Foundations of Ecology. Classic Papers with Commentaries. In this very important paper, MacArthur and Pianka developed a graphical model of animal feeding activities based on costs versus profits. A forager's optimal diet can be specified and some interesting predictions emerge. Prey abundance influences the degree to which a consumer can afford to be selective because if affects search time per item eaten. Diets should be broad when prey are scarce (long search time), but narrow if food is abundant (short search time) because a consumer can afford to bypass inferior prey only when there is a reasonably high probability of encountering a superior item in the time it would have taken to capture and handle the previous one. Also larger patches should be used in a more specialized way than smaller patches because travel time between patches (per item eaten) is lower. Behavioral ecologists quickly embraced OFT because it conferred apparent rigor and generated testable predictions in a sometimes subjective field. Theoreticians explored a variety of ways to employ OFT, which proliferated to become a minor growth industry during the 1970s and early 1980s. Studies of foraging behavior have proliferated extensively over the past three decades and several books have been written about foraging. Pianka (1966) also adopted a more descriptive empirical approach based on his observations of desert lizards, identifying two distinct modes of foraging: sit-and-wait (ambush predators) versus widely-foraging (more active predators). Of course, this dichotomy could be somewhat artificial, although numerous animal groups seem to fall rather naturally into either one category or the other. Members of most lizard families typically exploit either one or the other of these two modes of foraging: thus iguanians, agamids and geckos primarily sit-and-wait for their prey, whereas teiids and skinks forage widely. Lacertids, however, exploit both modes of foraging, even within the same genus. This evidently natural dichotomy in foraging tactics has had a substantial impact on theories of optimal diets and competitive relationships among species. Certain dietary differences are associated with this apparent dichotomy in foraging tactics. Sit-and-wait predators rely largely on moving prey whereas widely-foraging predators encounter and consume non-moving types of prey items more frequently. For the sit-and-wait tactic to pay off, prey must be relatively mobile and prey density must be high (or predator energy requirements low). The sit-and-wait tactic should be less prevalent during periods of prey scarcity than the widely-foraging method. The success of the widely-foraging tactic is also influenced by prey mobility and prey density as well as by the predator's energetic requirements (which should usually be higher than those of sit-and-wait predators), but the searching abilities of the predator and the spatial distribution of its prey now assume substantial importance. Another important spin-off of foraging mode involves reproductive tactics. Clutch sizes of widely-foraging species are smaller than those of sit-and-wait species, probably because the former simply cannot afford to weight themselves down with eggs to as great an extent as can the latter. Hence foraging style constrains reproductive prospects (as well as vice versa). Huey and Pianka (1981) summarized many of these ecological correlates of foraging mode. So-called optimal foraging theory and Pianka's modes of foraging constitute two distinct approaches to foraging that have evolved and begun to coalesce during the past decade. The literature on animal foraging was reviewed in Trends in Ecology and Evolution by Perry and Pianka (1997), and elevated to a paradigm in a 2007 book Lizard Ecology: The Evolutionary Consequences of Foraging Mode (Reilly, L. D. McBrayer, and D. Miles, eds.) with an introduction by Huey and Pianka and a chapter by Vitt and Pianka. The emerging new framework is more complex than originally thought, combining both theory and observation. Modern phylogenetic methods promise powerful new insights into how animal foraging has evolved. Natural selection in saturated environments (where demand for resources approximates supply) is density dependent, favoring competitive ability at the expense of slow growth and delayed reproduction. In contrast, in competitive vacuums (resource supplies greatly exceed demand), selection is independent of population density and favors rapid growth, early reproduction, and short life spans. Concepts of r and K selection were clarified and their correlates listed by Pianka (1970). This paper was reprinted in 1978 in Readings in Sociobiology and was chosen as a Citation Classic in 1979. It is usually cited in textbooks and papers on life history tactics and its summary table has been reprinted many times. A sequel to this paper (Pianka 1976), entitled Natural Selection of Optimal Reproductive Tactics, is an equally important general contribution although it is not cited as much. Selection recognizes only one currency: successful offspring. Yet even though all living organisms have presumably been selected to maximize their own lifetime reproductive success, they vary greatly in exact modes of reproduction. High fecundity early in life is usually correlated with decreased fertility later on. Plots of fecundity versus age are typically much flatter when early fecundity is lower. (This is an excellent example of the principle of allocation.) Some organisms, such as most annual plants, a multitude of insects, and certain fish like the Pacific salmon, reproduce only once during their entire lifetime. These big-bang or semelparous reproducers typically exert a tremendous effort in this one and only opportunity to reproduce (in fact their exceedingly high investment in reproduction may well contribute substantially to their own demise!). Many other organisms, including perennial plants and most vertebrates, do not engage in such suicidal bouts of reproduction but reproduce again and again during their lifetime. Such organisms have been alled iteroparous (repeated parenthood). Even within organisms that use either the big-bang or the iteroparous tactic, individuals and species differ greatly in numbers of progeny produced. Annual seed set of different species of trees ranges from a few hundred or a few thousand in many oaks (which produce relatively large seeds) to literally millions in redwood trees. Seed production may vary greatly even among individual plants of the same species grown under different environmental conditions; an individual poppy produces as few as four seeds under stress conditions, but as many as a third of a million seeds hen grown under conditions of high fertility. Fecundity is equally variable among fish; the ocean sunfish, Mola mola, is perhaps the most fecund of all vertebrates with a clutch of about 200 million tiny eggs. A female codfish also lays millions of relatively tiny eggs. Most elasmobranchs (sharks, skates and rays), however, produce considerably fewer but much larger offspring. Variability of clutch and/or litter size is not nearly so great among other classes of vertebrates, but it is still significant. Among lizards, for example, clutch size varies from a fixed clutch of one in some geckos and Anolis to as many as 40 in certain horned lizards (Phrynosoma) and the large Iguana. Timing of reproduction also varies considerably among organisms. Due to the finite chance of death, earlier reproduction is always advantageous, all else being equal. Nevertheless, many organisms postpone reproduction. The century plant, an Agave, devotes years to vegetative growth before suddenly sending up its inflorescence (some related monocots bloom much sooner). Delayed reproduction also occurs in most perennial plants, many fish such as salmon, a few insects like cicadas, some lizards, and many mammals and birds, especially among large seabirds. Innumerable other examples of the diversity of existing reproductive tactics could be listed. Clearly, natural selection has shaped observed reproductive tactics, with each presumably corresponding in some way to a local optimum that maximizes an individualís lifetime reproductive success in its particular environment. Population biologists would like to understand factors that influence evolution of various modes of reproduction. How much should an organism invest in any given act of reproduction? R. A. Fisher (1930) anticipated this question over 50 years ago:
Fisher distinguished clearly between the proximate factor (physiological mechanism) and the ultimate factors (circumstances in the life history and environment) that determine the allocation of resources into reproductive versus non-reproductive tissues and activities. Loosely defined as an organism's investment in any current act of reproduction, reproductive effort has played a central role in thinking about reproductive tactics. Although reproductive effort is conceptually quite useful, it has yet to be adequately quantified (Pianka 1976). Ideally, an operational measure of reproductive effort would include not only the direct material and energetic costs of reproduction but also risks associated with a given level of current reproduction. Another difficulty concerns the temporal patterns of collection and expenditure of matter and energy. Many organisms gather and store materials and energy during time periods that are unfavorable for successful reproduction but then expend these same resources on reproduction during a later, more suitable time. The large first clutch of a fat female lizard that has just over-wintered may actually represent a smaller investment in reproduction than her subsequent smaller clutches that must be produced with considerably diminished energy reserves. Reproductive effort could perhaps be best measured operationally in terms of the effects of various current levels of reproduction upon future reproductive success. In spite of these rather severe difficulties, instantaneous ratios of reproductive tissues over total body tissue are sometimes used as a crude first approximation of an organism's reproductive effort (both weights and calories have been used). Thus measured, the proportion of the total resources available to an organism that is allocated to reproduction varies widely among organisms. Among lizards, relative clutch mass (the fraction of a gravid female's mass devoted to progeny) varies from 4-5% to 40-50% (Pianka 1986). Among different species of plants, energy expenditure on reproduction, integrated over a plant's lifetime, ranges from near zero to as much as 40 percent. Annual plants tend to expend more energy on reproduction than most perennials (about 14 to 30 percent versus 1 to 24 percent). An experimental study of an annual euphorb showed that calories allocated to reproduction varied directly with nutrient availability and inversely with plant density and competition. Returning to Fisher's dichotomy for the apportionment of energy into reproductive versus non-reproductive (somatic) tissues, organs, and activities, Pianka and Parker (1975) and Pianka (1976) worked out the theory of optimal reproductive effort. Somatic tissues are clearly necessary for acquisition of matter and energy; at the same time, an organism's soma is of no selective value except inasmuch as it contributes to that organism's lifelong production of successful offspring. Allocation of time, energy, and materials to reproduction in itself usually decreases growth of somatic tissues and often reduces future fecundity. Increased reproductive effort may also cost by reducing survivorship of the soma; this is easily seen in the extreme case of big-bang reproduction, in which the organism puts everything available into one suicidal bout of reproduction and then dies. More subtle changes in survivorship also occur with minor alterations in reproductive effort. How great a risk should an optimal organism take with its soma in any given act of reproduction? To explore this question, Pianka (1976) exploited the concept of residual reproductive value, which is simply age-specific expectation of all future offspring beyond those immediately at stake. Residual reproductive value thus measures the present value of the soma. To maximize its overall lifetime contribution to future generations, an optimal organism should weigh the profits of its immediate prospects of reproductive success against the costs to its long-term future prospects. An individual with a high probability of future reproductive success should be more hesitant to risk its soma in present reproductive activities than another individual with a lower probability of reproducing successfully in the future. Moreover, to the extent that current reproduction decreases expectation of further life, it may reduce residual reproductive value directly. For both reasons, current investment in reproduction should vary inversely with expectation of future offspring. The inverse interaction between reproductive effort and residual reproductive value can take several possible different forms. Pianka (1976) developed a simple graphical model that relates costs and profits in future offspring, respectively, to profits and costs associated with various levels of current reproduction, the latter measured in present progeny. Possible tactics available to a given organism at a particular instant range from a current reproductive effort of zero to all-out big-bang reproduction. An optimal reproductive tactic exists for any given set of possible tactics when lifetime reproductive success is maximized. 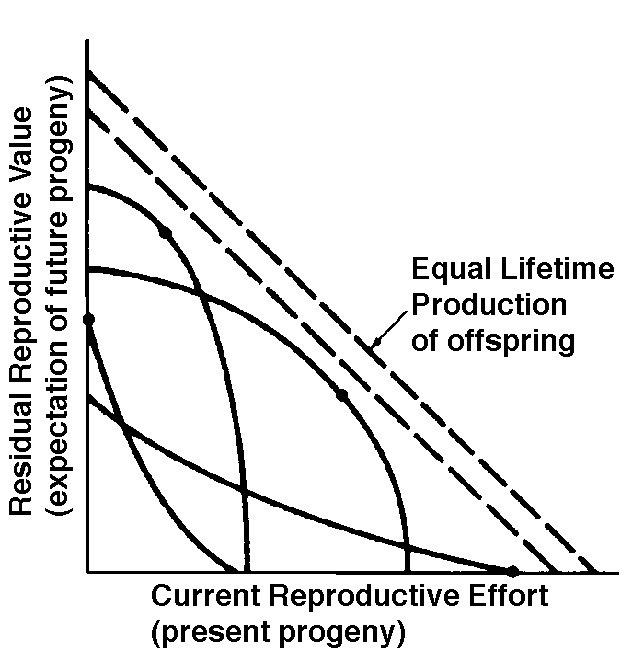 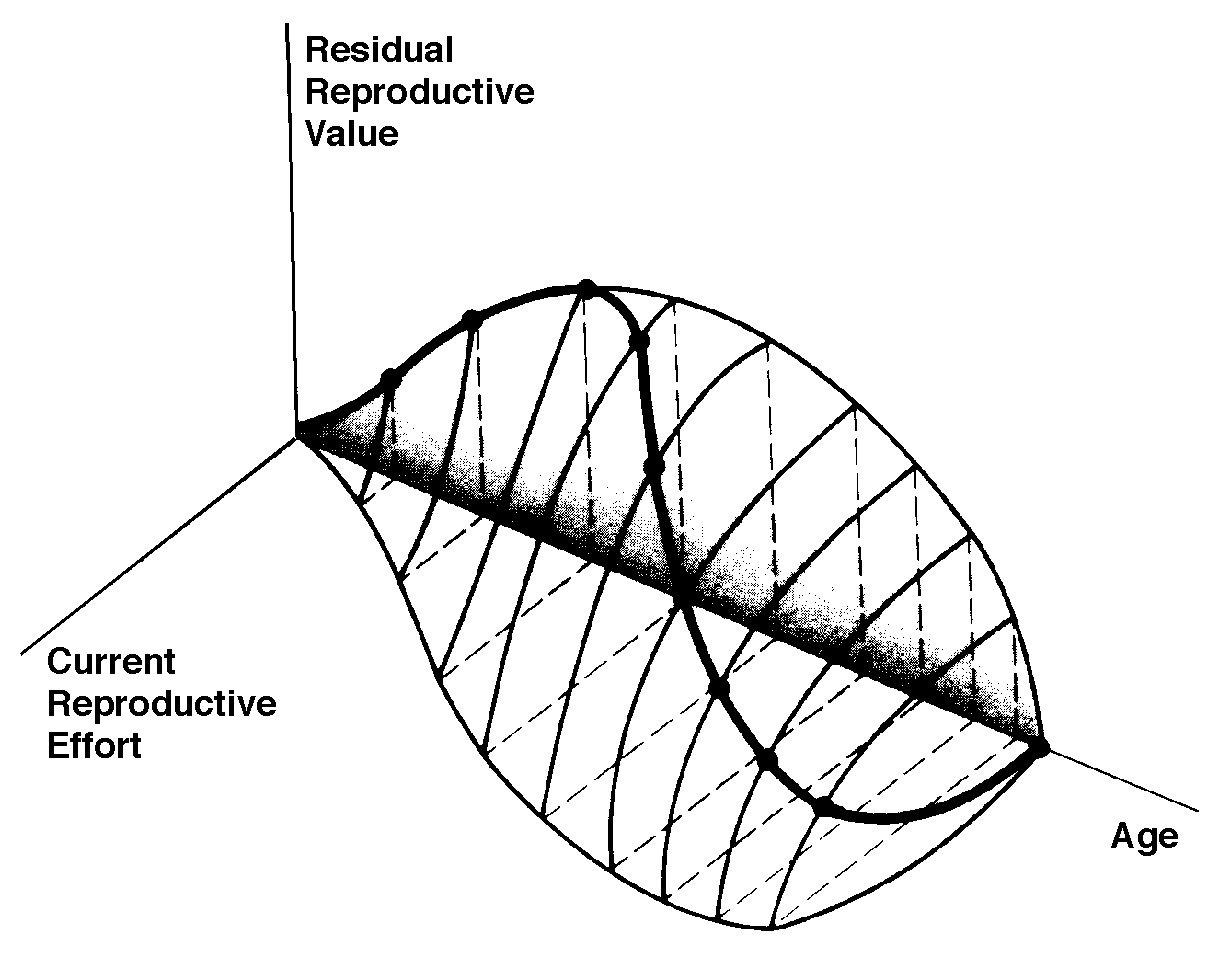 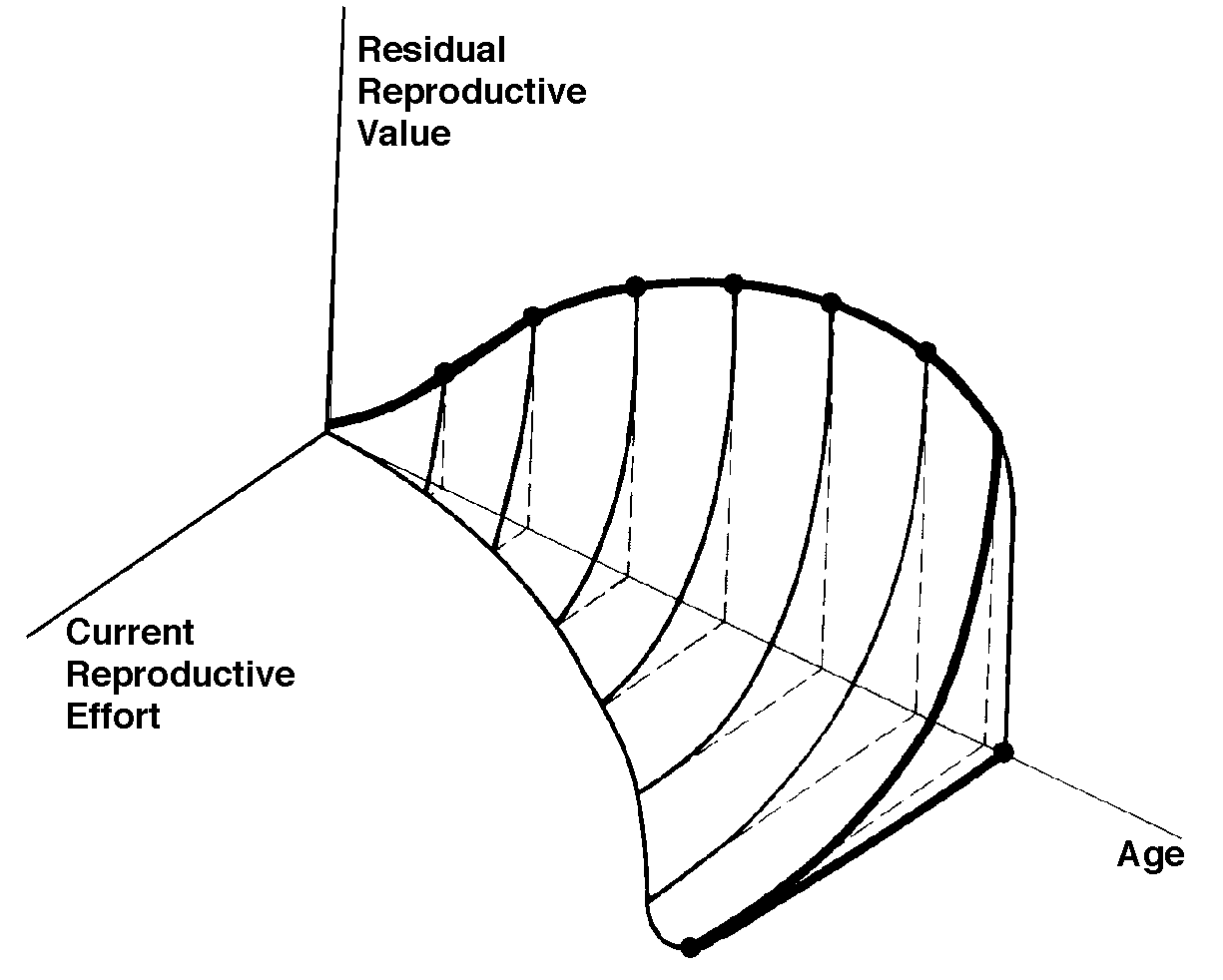
The exact form of tradeoffs between present and future reproduction depend on the actual reproductive tactic taken by an organism as well as on immediate environmental conditions for foraging, reproduction, and survival. Numerous factors influence the precise form of these trade-offs, including predator abundance, resource availability, and numerous aspects of the physical environment. Unfavorable conditions for immediate reproduction decrease costs of allocating resources to somatic tissues and activities, resulting in lower reproductive effort. (Improved conditions for survivorship, such as good physical conditions or a decrease in predator abundance, would have a similar effect by increasing returns expected from investment in soma.) Conversely, good conditions for reproduction and/or poor conditions for survivorship result in greater current reproductive effort and decreased future reproductive success. Pianka (1976) also worked out a theory for optimal expenditure per progeny, as follows. Not all offspring are equivalent. Progeny produced late in a growing season often have lower probabilities of reaching adulthood than those produced earlier -- hence, they contribute less to enhancing parental fitness. Likewise, larger offspring may usually cost more to produce, but they are also worth more. How much should a parent devote to any single progeny? For a fixed amount of reproductive effort, average fitness of individual progeny varies inversely with the total number produced. One extreme would be to invest everything in a single very large but extremely fit progeny. Another extreme would be to maximize the total number of offspring produced by devoting a minimal possible amount to each. Parental fitness is often maximized by producing an intermediate number of offspring of intermediate fitness: Here, the best reproductive tactic is a compromise between conflicting demands for production of the largest possible total number of progeny (r selection) and production of offspring of the highest possible individual fitness (K selection). The optimal parental tactic is seldom the optimum for individual offspring, which achieve maximal fitness when parents invest everything in a single offspring. Hence, a parent-offspring conflict exists. The relationship between progeny fitness to parental expenditure in real organisms is influenced by a virtual plethora of environmental variables, including length of life, body size, survivorship of adults and juveniles, population density, and spatial and temporal patterns of resource availability. The competitive environment of juveniles is likely to be of particular importance because larger, better-endowed offspring should usually enjoy higher survivorship and generally be better competitors than smaller ones. Juveniles and adults are often subjected to very different selective pressures. Reproductive effort should reflect environmental factors operating upon adults, whereas expenditure per progeny will be strongly influenced by the environments encountered by juveniles. Because any two parties of this triumvirate determine the third, an optimal clutch or litter size is a direct consequence of an optimum current reproductive effort coupled with an optimal expenditure per progeny (indeed, clutch size is equal to reproductive effort divided by expenditure per progeny). Of course, clutch size can be directly affected by natural selection as well. Horned lizards, for example, are long-lived and relatively K-selected as adults, but because of their tank-like body form, these lizards have a very large reproductive effort and produce many tiny offspring which suffer very high mortality. Pianka (1973) pointed out that lizards partition environmental resources in three major ways: by being active at different times, by spending time in different places, and by eating different foods. These three niche dimensions are not independent, but together they serve to separate most pairs of sympatric species. Pianka (1973) also proposed a new symmetric index for quantifying niche overlap. Pianka was invited to review competition and niche theory for two editions of an important and influential book Theoretical Ecology edited by R. M. May (1976, 1981), which was translated into German. Pianka's 1974 PNAS paper entitled niche overlap and diffuse competition was reprinted in Niche: Theory and Application. Benchmark Papers in Ecology. He published several important articles on niche segregation (Pianka 1969, 1973; Pianka and Huey 1978; Pianka, Huey, and Lawlor 1979). Pianka has been an avid student of community structure for many years. He published an important paper on guild structure (Pianka 1980) and another on indirect effects (Pianka 1987). Pianka was invited to give the plenary lecture on The State of the Art of Community Ecology at the First World Congress of Herpetology in 1989 (published in 1992). He and his ex-graduate student Kirk Winemiller developed protocols for detecting and comparing the organization of different communities, using neotropical fishes and desert lizards as examples (Winemiller and Pianka 1990). Pianka's first paper on species diversity, a synthetic review of concepts behind latitudinal gradients in diversity (Pianka 1966), was reprinted 15 years later in 1981 in Tropical Ecology. Benchmark Papers in Ecology. This classic paper has strongly influenced this important field ever since. He wrote a chapter on reptile species diversity (Pianka 1977) plus several papers on lizard diversity (Pianka 1967, 1981, 1989, 1994). In addition, he published two more general chapters on species diversity (Pianka 1971, 1979). Pianka's work on patterns of species diversity is at the cutting edge in the developing field of biodiversity -- a subject receiving more and more attention as the planet's biota continues to disappear due primarily to human-induced loss of habitat. While many people talk a lot about diversity, no-one has dedicated themselves to its study to the extent that Pianka has. He is the consummate field biologist, but one who also appreciates and contributes to theory. His own personal long-term panglobal field studies of factors influencing desert lizard diversity are now widely recognized as a modern-day classic and are described in most ecology textbooks. The importance of spatial scale has been neglected in traditional ecology, although not in the emerging field of landscape ecology. While the implications of the landscape on ecology have long been appreciated, only recently have quantitative methods of study been exploited. In the past, ecologists, including Pianka, have focused on local-level processes. Larger scale regional factors also control local phenomena. Local species richness may often be a consequence of regional processes. Relatively little empirical attention has been given to the interaction between these two levels. Unfortunately, few complete closed regions remain unfragmented by human activities in which regional and local phenomena can still be studied simultaneously. Pianka has undertaken such a study in the uninhabited Great Victoria desert of Western Australia, an area with an extremely high diversity of lizards. Fires were once a major agent of disturbance in all grassland and semi-desert biomes, including the North American tall grass prairies. Most of these ecosystems have now been reduced to mere vestiges, and controlled burning and/or fire control are practiced by humans almost everywhere. The inland Australian desert is one of the last remaining areas where natural wildfires remain a regular and dominant feature of an extensive natural landscape largely undisturbed by humans. In this region, an important fire succession cycle generates spatial and temporal heterogeneity in microhabitats and habitats. Habitat-specialized species can go extinct within a given habitat patch (fire scar), but remain in the overall system by periodic reinvasions from adjacent or nearby patches of suitable habitat of different age. These regional processes facilitate local diversity. Pianka has studied this system at the local level in the field in Western Australia and at the regional level at the University of Texas at Austin using aerial photography and multispectral satellite imagery. Macrographs, high resolution satellite images, offer a powerful new way, heretofore underutilized by biologists, to acquire regional level data on the frequency and phenomenology of wild fires, and thus the system-wide spatial-temporal pattern of disturbance. Several different approaches to modelling fire succession are being examined. Fire spread was modelled, both analytically and using computer simulation. These include a fractal-based model, a Poisson random field model, a cellular automata model, and a Markov random field model, all of which use empirical transition probabilities. Dozens of Landsat Multispectral Scanner (MSS) satellite images of two scenes (which includes two long-term study sites) were acquired over the 22-year interval from 1972 to 1994, including all years from 1979-88 plus some expensive more recent imagery. All scenes are cloud free and were accessed during October or November. Images were destriped, radiometrically corrected, and georeferenced (geometrically registered) to one another by stapling images together at identifiable known ground control points and rubber sheeting. High resolution Landsat Thematic Mapper (TM) and SPOT imagery for this region were acquired and registered to the MSS imagery and used for ground truthing, along with high altitude radar and low altitude aerial photography. NASA flew an AIRSAR overflight over our study region in September 1993. Ground level radiometric measurements, additional ground truth, wind and precipitation data, and GPS coordinate data were acquired. Much of the digital satellite data has been acquired by Landsat, archived on magnetic tape. A complete analysis of wild fires for the Great Victoria desert region required over a hundred images. Imagery was purchased and analyzed to detect burned areas. Spectral and spatial statistics were computed for over a thousand fires, and the probability that a given area will burn was estimated. Other data collated for each fire include: date, location, area, perimeter, compass direction (of burn and prevailing wind), ground cover characteristics, extent of reticulation, as well as various fractal dimensions. Age and size distributions of burn patches were estimated. Eventually, Pianka hopes to acquire supporting imagery from other grassland areas, particularly the Kalahari semidesert of southern Africa, to use for comparative purposes (fires in the Kalahari do not appear to reticulate to as great an extent as they do in Western Australia). A major goal of this study is to obtain baseline data on temporal patterns, spatial structure, and distribution of disturbances. These data will form the backdrop for a more detailed study of the population dynamics and dispersal abilities of species. To read more about this project, Click here. Fire scars are quite evident and are easily followed through time. Several hundred fire scars were extracted from the imagery and incorporated into a GIS. Changes in reflection patterns through time, subtraction and classification analyses allowed discrimination of new burns, young spinifex, mature spinifex, marble gums, mallee, sandridges, mulga, salt lakes as well as rocky areas. Transition probabilities were estimated from chronosequences of satellite images. Geology and vegetation maps for the area were digitized and extensive rainfall and wind data were acquired and input to the GIS for modelling efforts still in progress. The landscape is a mosaic of habitat patches of varying sizes that is constantly changing. Habitat-specialized species can go extinct within a given habitat patch (fire scar), but remain in the overall system by periodic reinvasions from adjacent or nearby patches of suitable habitat of different age. Such regional processes facilitate local diversity. Life in terrestrial Australian ecosystems has evolved over the past 10 million years to thrive in habitats kept in a dynamic state through fire succession cycles. Previous studies support the notion that wildfires promote species diversity in plant and animal communities by creating a heterogeneous mix of habitats, each habitat more suitable for particular subsets of species. A new study site with pit trap lines was set up and monitored for several years before a controlled burn. Pianka performed a controlled burn of this site, the B-area, in early October 1995. He monitored this study site over the next two decades to assess faunistic changes (especially among lizards, but also arthropods eaten by lizards) following fire. Pianka and his graduate student Stephen Goodyear documented population and community responses to fire in a species-rich lizard assemblage in the Great Victoria Desert of Western Australia. Lizards were censused by pit trapping at a long-unburned flat spinifex site in the Great Victoria Desert in Austral springs of 1992 and 1995. A controlled burn was undertaken in mid-October of 1995, and lizards were censused thereafter in late 1995 and early 1996, and then again in the Austral springs of 1998, 2003, and 2008. Forty-six species of lizards (2872 individuals) were collected and their stomach contents analyzed over the course of a 16-year fire succession cycle at this single study site. Most strikingly, relative abundances of two species of agamids varied inversely, responding oppositely to habitat clearing effects of fire. The military dragon Ctenophorus isolepis reached higher abundances when vegetation was dense, and decreased in abundance in open vegetation following fire. The netted dragon Ctenophorus nuchalis was rare when vegetation coverage was high but increased rapidly after fire. Abundances of five species of Ctenotus skinks, C. ariadnae, C. calurus, C. hanloni, C. pantherinus, and C. piankai, tracked those of Ctenophorus isolepis. Abundance of a termite-specialized nocturnal gecko, Rhynchoedura ornata, increased in abundance following fire. Lizard diets changed during the course of the fire succession cycle, returning to near pre-burn conditions after 16 years. In addition to short-term fire succession cycles that contribute to structuring local communities, changes in long-term rainfall also impact desert food webs and regional biotas. Inevitably, environments that support a diverse assemblage of closely-related species are sub-divided into a number of different microhabitats. Complex environments sub-divided into many such microhabitats support more species than simpler environments. Such habitat heterogeneity implies a decreased absolute extent of any one habitat type, existence of habitat edge, plus a degree of fragmentation -- all of which are usually associated with a detrimental impact on the persistence of individual species. An intriguing paradox arises: the existence of a mosaic of habitat patches is critical to the maintenance of species diversity in the environment as a whole, yet the resulting fragmentation potentially threatens the persistence of every population of the constituent species within the coexisting assemblage. Clearly, maintenance of the appropriate spatial scale of such habitat mosaics is critical to their ability to support diverse multi-species assemblages. Moreover, when considering habitats structured and characterized by different vegetation types, their spatial configuration is likely to vary temporally, generating a complex mosaic of transient habitat patches, the dynamics of which are driven by some successional process. The ability of individual species to persist in such spatially and temporally changing environments depends on their ability to track (locate and exploit) suitable habitat as and when it occurs. The relative connectivity of similar patches, and the permeability to the inhabiting species of the landscape as a whole is therefore also of critical importance, suggesting that the shape of patches and the texture of their agglomerate is also directly relevant to long-term survival of the assemblage. Animals with active habitat selection such as Australian desert lizards should reach ecological and evolutionarily stable equilibria between source and sink habitats, with dispersal from the former to the latter maintaining the species locally. Such data on the metapopulations of component species will eventually be fitted into the overall spatial-temporal mosaic in an effort to explain the persistence of this diverse desert fauna. These lizard populations can be viewed more realistically using a metaphor: relative abundances are analogous to 3-D waves in as many dimensions as there are species: the vertical coordinate represents abundances of each of the various species moving up and down and around in space and time. At any given time, some species are abundant while others may be scarce. These abundance surfaces are relatively placid for some species, but very rough for others. Some species like Ctenophorus clayi and Ctenophorus nuchalis boom and bust, exhibiting intermittent rarity, whereas others like Cyclodomorphus, Eremiascincus, and Tiliqua, are always uncommon (chronic rarity). Still other abundant species are common, like Ctenophorus isolepis, with more stable populations. These waves of relative abundance respond to fire and precipitation, both of which drive changes in resource availabilities of prey and microhabitats through time and space (fire succession cycle). One of Pianka's long-term goals is to attempt to describe this multidimensional spatial-temporal wave-like landscape. Ultimately, Pianka plans to model the entire Great Victoria desert region as a dynamic habitat mosaic so as to understand mechanisms of coexistence of its component species and the effects of fire disturbance in maintaining diversity in this region. Faunas of unstudied sites will be predicted by extrapolation, thus allowing models to be to subjected to direct test. It should also be feasible to exploit controlled burns to test some aspects of such fire succession models. Geographic variation in the number of coexisting plant and animal species (species density) often follows repeated patterns; best known is the general increase in species richness from temperate to tropical latitudes. With his ex-graduate student Jos. J. Schall, Pianka undertook a detailed intercontinental comparison of geographical trends in species richness (Schall and Pianka 1977, 1978, Pianka and Schall 1981). Their major paper was published as the lead article in Science (Schall and Pianka 1978). They performed an elaborate multivariate quantitative analysis of the numbers of species of terrestrial vertebrates (amphibians, reptiles, birds, and mammals, total number of species about 2500) of the United States and Australia. Trends in numbers of species were described and correlated with geographic variation in abiotic environmental measures. Latitudinal gradients in species density were strong in North America, but were not evident in Australia. Correlations between species densities and abiotic environmental factors for birds and mammals differ on the two continents: correlations are often opposite in sign. Intercontinental comparisons revealed general patterns as well as some intriguing and profound differences in vertebrate distributions. For example, only about 100 species of lizards occur in the continental U.S.A. The Australian continent has about the same land area, but supports 400-500 lizard species, four to five times as many species. On both continents, lizards have diversified most in arid sunny regions, probably partially because costs of thermoregulation are low for ectotherms in such environments. The ability to become inactive during periods of stress confers ectotherms with an advantage over endotherms in unpredictable desert environments. Australian lizards are also much more diverse ecologically than are North American lizards. How have lizards undergone such extensive speciation so rapidly within continental Australia? Relatively few obvious geographic barriers, such as mountains, lakes, and/or rivers, exist in the Australian desert region. Zones of unsuitable habitat form the most effective barriers to lizard dispersal. The high degree of habitat specificity observed in Australian desert lizards, coupled with extensive spatial patchiness in the mosaic of desert vegetation, led Pianka to propose a simple model for lizard speciation based on fluctuating boundaries among three major habitat types to which Australian desert lizards have evolved habitat specificity (Pianka 1969a, 1972). Some species of Australian desert lizards are restricted to flat spinifex sandplains, others to shrub-Acacia (mulga) habitats, and still others to sandridges. Habitat-specific lizard species, such as those restricted to shrub-Acacia (mulga) habitats, move along appropriate corridors. Long-term changes in climate and/or soils, such as movements of windblown sands, could easily break shrub corridors, separating eastern from western patches of shrub-Acacia habitats and isolating shrub-specific lizards to diverge and speciate. Simultaneously, sand corridors are opened, allowing sand-specialized stocks to invade previously unoccupied sandy desert regions. Later in geological time, shifting sands could reverse the process, resulting in closure of sand corridors, forming isolates among sand species while reopening shrub corridors. Such alternating habitat junctures probably isolated eastern from western populations of lizards restricted to shrub-Acacia habitats as well as northern from southern (in central and western Australia) and eastern from western populations of species restricted to sandridge and/or sandplain-Triodia habitats (see maps in Pianka 1972). In support of the above hypothesis, the sandridge-specialized Ctenotus brooksi species complex has several recognized taxa, euclae in the coastal sand dunes along the south coast, iridis in the Victorian Little desert, taeniatus on the sands surrounding Lake Torrens, aranda in the Simpson desert, and brooksi in the Great Victoria and Gibson deserts. Knowledge of phyletic relationships of these taxa will provide exceedingly valuable insights into vicariant events that took place during the history of arid Australia. A number of putatively closely-related species pairs also live together in the same habitat but have evolved other niche differences, such as Ctenotus colletti and leae, as well as piankai and quattuordecimlineatus. Do closely-related sympatric species diverge in ecology more than distantly-related species? With a phylogeny, one can test whether or not various assemblages actually known to coexist in various geographic regions or at particular study sites are either more or less closely-related than expected by comparing their phylogenetic distances against sets of species drawn at random in repeated Monte-Carlo simulations (Pimm 1983, Gotelli and Graves 1996). Ecological similarities will also be compared with phylogenetic distances (Haydon et al. 1993). Members of some Ctenotus species groups, such as the putative C. colletti and C. schomburgkii groups, contain species that have become specialized to different habitat elements, suggesting that habitat specificity has evolved (brooksi, colletti and leae are sandridge species, whereas putatively closely-related calurus and schomburgkii are denizens of sandplain-Triodia habitats. In the putative atlas species group, dux is a sandridge species, whereas piankai and quattuordecimlineatus are sandplain species. It will be most instructive to overlay habitat preferences on a resolved phylogeny to determine the extent of evolutionary lability in habitat specificity. In support of Lack's (1968) hypothesis, Richman and Price (1992) found that habitat difference evolved last in Palearctic leaf warblers. Will the same conclusion hold true for these desert lizards? The time frame for the evolution of the genus Ctenotus is closely comparable to current estimates of the period of aridification of the Australian continent. Have Ctenotus radiated in response to the progressive enlargement and diversification of the arid zone? If so, then a biogeographic analysis of Ctenotus, undertaken within a phylogenetic framework, should reveal a wealth of information about the process and history of aridification itself. Pertinent questions include: 1) Are arid-adapted Ctenotus derived from within species groups of semiarid and temperate origin, or is the reverse true? 2) Have various habitat islands of hard country played a role in the evolution of Ctenotus? Six other Australian squamate genera are also species-rich: [Lerista (about 80 spp.), Diplodactylus (36 spp.), Egernia (28 spp.), Varanus (25 spp.), Ctenophorus (21 spp.), Ramphotyphlops (31 spp.)] -- these need to be examined in a comparative sense. Ctenotus, the most species rich and most widespread genus, represents the ideal test case for development of methods and will provide an incentive for equivalent studies in other groups. Obviously, congruence will lend credence to hypothesized vicariant events. Such studies will also supply the context for predictions regarding diversity of other groups of arid zone animals/plants and will have obvious management implications, especially with regard to conservation of genetic diversity. Deduction of biogeographic process from the study of history is complicated by the fact that history is singular. This singularity allows no estimation of degree of determinism surrounding the realization of actual biogeographic processes. Consequently, almost nothing is known about the generality of such deductions. The current diversity and distribution of Earth's biota constitute but a single snapshot -- one possible realization of a vast and ongoing probability experiment. The particular realization observed has generated a variety of apparently non-random patterns (latitudinal gradients in species diversity, to name just one). Such biogeographic patterns result from a long-term, auto-contingent sequence of dispersal, speciation, and local extinction taking place over a continuous series of tectonic events. Haydon, Radtkey and Pianka (1993) and Haydon, Crother and Pianka (1994) developed a new approach, called experimental biogeography, that exploits computers to simulate faunal buildup repeatedly against a fixed vicariant background over ecological and evolutionary time scales. This new approach enables a biogeographer to be omniscient -- to know both vicariant history and actual phylogeny. Moreover, history can be replayed repeatedly to accumulate a sample of multiple phylogenies and to estimate probability density functions for biogeographic variables. Roles of stochastic, historical and ecological processes in adaptive radiations can also be assessed. Experimental biogeography allows examination of the reliability of various methods of recovering historical patterns. Over the last couple decades, evolutionary biologists have become acutely aware that, due to shared common ancestry, related species do not constitute independent observations (Felsenstein 1985). To factor out phylogenetic effects requires recovering the phylogeny of that fauna (Ridley 1983; Brooks and McLennan 1991). Appropriate statistical procedures to do this were first worked out by Ridley (1983) and Felsenstein (1985, 1988), and extended by Grafen (1989), Maddison (1990), Harvey and Pagel (1991), Martins and Garland (1991), Garland (1992), Garland et al. (1992, 1993), Hansen and Martins (1996), Martins (1996), Martins and Hansen (1997), among others. Some of the greatest potential benefits of the recent emphasis on phylogeny may accrue in the area of ecology (Brooks 1985; Futuyma and Moreno 1988; Brooks and McLennan 1991; Harvey 1991), but ecological applications using phylogenetic data are unfortunately scant. Losos (1990a) examined the influence of phylogeny on body size and character displacement of pairs of species in the Caribbean lizard genus Anolis and suggested that, although size evolution is rare, similar-sized species in sympatry evolve in different directions (size adjustment, or character displacement), presumably reducing interspecific competition. Habitat difference evolved last in Palearctic leaf warblers (Richman and Price 1992). In the fish genus Cyprinella, species branching near the cladogram root had larger geographical ranges than those branching distally (Taylor and Gotelli 1994). Other groups need to be studied to determine the generalizability of such results. Pianka is currently undertaking a phylogenetic analysis of the geography and adaptive radiation of Australian skinks of the genus Ctenotus. Anatomical and ecological convergences can be identified by simply plotting character transformations on a phylogeny. Similarly, with a known phylogeny, the evolution of niche transitions can be traced and probable sequences of evolutionary changes can be established. Covariant patterns of character change can be scrutinized to ascertain the extent to which phylogeny has constrained morphology and ecology. Modern comparative methods allow examination of the probable course of evolution in a lineage of lizards (family Varanidae, genus Varanus). Within this genus, body mass varies by nearly a full five orders of magnitude. The fossil record and present geographic distribution suggest that varanids arose over 65 million years ago in Laurasia and subsequently dispersed to Africa and Australia. Two major lineages have undergone extensive adaptive radiations within Australia: one evolved dwarfism (subgenus Odatria = pygmy monitors), whereas the other Australian lineage (subgenus Varanus) remained large and several of its members evolved gigantism. Pianka (1995, 2005) plotted body sizes of extant varanid species on a phylogeny and inferred probable sizes at ancestral nodes from those of their descendents. Phylogenetically independent contrasts, coupled with information on branch lengths, were exploited to identify several likely instances of relatively rapid evolution of body size, both between and within clades. Pianka discussed possible scenarios for evolution of size in this lizard lineage and raised many questions worthy of further research. Numerous questions about the evolution of size in this genus within a historical/geographical perspective remain to be answered. Varanus are morphologically conservative, but vary in mass by nearly five orders of magnitude. No other terrestrial animal genus exhibits such a range of size variation. There is proportionately almost as much difference in mass among species within the genus Varanus as there is between an elephant and a mouse [Mammals vary in size from 3 gm shrews to whales that weigh over 50 tons, but constitute an entire subclass. No mammal genus exhibits size variation anywhere near as extensive as Varanus. Varanids are sufficiently conservative morphologically that taxonomic ranking at the generic level seems justified. Even if the genus Varanus is eventually broken up into several closely-related genera, the comparison with size variation in mammals is still striking.] These properties, coupled with the fact that a phylogeny has recently become available, make varanid lizards a nearly ideal model system for examining patterns of size evolution. All living members of the Varanidae, commonly known as monitor lizards, are placed in a single genus Varanus, generally considered to be monophyletic (Baverstock et al. 1993; Green and King 1993; King et al. 1991; Pregill et al. 1986). Many monitor species are large and impressive -- they are often the centerpiece of reptile house exhibits. Varanus, which are not particularly tractable research subjects, have also received an extraordinary amount of attention from dedicated students. Except for a small clade of frugivorous Philippine monitors, all are active predators and eat quite large prey relative to their own body size (Pianka 1982, 1994, 2005; but see Losos and Greene 1988 and Auffenberg 1994 for dissenting views). Many monitor lizards are top predators (Losos and Greene 1988; Pianka 1994). Some species are aquatic, others terrestrial, while still others are saxicolous and/or semi-arboreal or truly arboreal. Monitor lizards live in a wide variety of habitats, ranging from mangrove swamps to dense forests to savannas to arid deserts. Two distinct evolutionary lineages have undergone extensive adaptive radiations in Australia: the pygmy monitor subgenus Odatria, and the subgenus Varanus, which includes the larger species of Australian monitors plus several species found nearby outside Australia (indicus, prasinus and teriae appear to have recently reached Australia from New Guinea). Australian monitors range in size from the diminutive pygmy monitor Varanus brevicauda (0.2 m in total length, 8-10 gms) to the largest living lizard in Australia, the perentie, Varanus giganteus (total length may exceed 2.0 m, mass 17 kg). The typical monitor lizard threat posture and behavior has been conserved in the evolution of tiny monitors such as brevicauda, which hiss and lunge with throat inflated as if they are a serious threat. Even more awesome are Komodo monitors (Varanus komodoensis) of the Lesser Sunda Islands of Indonesia, which attain total lengths of over 3 m and weights of more than 150 kg (Auffenberg 1981). Komodo monitors, however, are themselves dwarfed by the largest known terrestrial lizard, a closely-related gigantic varanid formerly assigned to the genus Varanus. Megalania prisca is a Pleistocene fossil (19,000-26,000 years BP) from Australia, estimated to have reached 7 m in total length and to have weighed more than 600 kg (Hecht 1975; Auffenberg 1981; Rich 1985). [Cladistic systematics will undoubtedly require that the fossil taxon Megalania eventually be reassigned to the genus and subgenus Varanus.] These spectacular creatures must have been even more formidable than modern-day saltwater crocodiles. The major prey of these gigantic monitor lizards is thought to have been large diprotodont marsupials (rhinoceros-sized beasts, now extinct, that were relatives of wombats and koalas). Being contemporaneous with aboriginal humans in Australia, in all probability, Megalania also ate Homo sapiens. Megalania teeth were over 2 cm long, curved, with the rear edge serrated for cutting and tearing the skin and flesh of its prey as these powerful predators pulled back on their bite. Many other species of Varanus also possess such teeth. Several authors have suggested that Varanus komodoensis and Megalania prisca are/were ecological equivalents of large saber-toothed cats, using a slashing bite to disembowl large mammals (Akersten 1985; Auffenberg1981; Losos and Greene 1988). Water buffalo as large as 590 kg have been killed by Varanus komodoensis, more than three times their own mass (Auffenberg 1981). Body size and clutch size have been plotted on varanid phylogenies (for examples, see Pianka 1995, 2005). The vast majority of genera contain only a few species. Most species lineages tend to exhibit niche conservatism, seldom changing much in evolutionary time. Species-rich genera are rare and extremely interesting because they imply that recent bouts of speciation and niche diversification have occurred, leading to an adaptive radiation. Such events of rapid evolution promote species diversity. However, major adaptive radiations are uncommon and few have been studied: those that have been include Hawaiian Drosophila, African Rift Lake cichlid fishes, Galapagos finches, and Anolis lizards. The Australian skincid genus Ctenotus has undergone a very extensive adaptive radiation within the last 4-10 million years. It is the most speciose lizard genus (about100 species) in Australia and among the most species rich lizard genera in the entire world. A phylogeny of Ctenotus has been recovered by sequencing DNA from both mitochondrial genes (12S, 16S, and ND4) and nuclear genes that are evolving at varying rates. Preliminary study of these genes has identified strongly supported clades and challenged the monophyly of several putative species groups. A resolved phylogeny of Ctenotus will allow tests of both the monophyly of the genus and current phenetic groupings within Ctenotus. Species groups will be redefined and, in all probability, new species will be discovered. Once a resolved phylogeny is in place, extensive ecological information will be examined from within a phylogenetic perspective to test various hypotheses about evolution and biogeography in this diverse genus. Traits that have evolved rapidly among species within this genus include body size, toe lamellae, body temperatures, habitat specificity, dietary and microhabitat niche breadth, head length, relative eye size, relative hindleg length, extent of geographic range, clutch size and other reproductive parameters such as relative clutch mass and size of oviductal eggs (expenditure per progeny). Anatomical and ecological convergences will be elucidated using phylogenetic systematics, modern comparative methods, and phylogenetically-based, multivariate statistical procedures. The extent to which phylogeny has constrained morphology and ecology will be ascertained. Biogeographic and phylogenetic studies will always be plagued by the fact that the past is singular, unknown, and we cannot return to it. Soft body parts, behavior and physiology do not fossilize. With information of the sort that will be gathered here, however, much can be inferred about past history -- probable regions of origin, speciation events, and dispersal events can be identified. Probable ancestral states will be inferred from those of surviving descendants. Historical biogeographic analysis will shed light on the origin and radiation of various species groups within Ctenotus, the aridification and vicariance of Australia, and will bear on the origin of the Australian arid zone fauna as a whole. Such a comprehensive study of a species rich genus will elucidate the factors that allowed this major adaptive radiation. It will also enable prediction of the probable ecologies of unstudied species of Ctenotus, which can then be tested by direct examination in the field. This study, now in progress, will elucidate factors 
allowing a major adaptiveradiation and will produce a phylogeny that will be used extensively. A series of papers will be published first in peer-reviewed journals and then an edited synthetic multiauthored volume with a University press. Publications will include papers and chapters on both (1) molecular and (2) morphologically based phylogenies, plus (3) a combined total evidence analysis, (4) a taxonomic rearrangement of species groups, (5) descriptions of new species (6) analyses of biogeography and geographic distributions, (7) evolution of microhabitat utilization and thermoregulation, (8) evolution and function of toe lamellae, (9) evolution of body size, (10) evolution of habitat preference, (11) evolution of diet, (12) evolution of reproductive tactics, and (13) an overview of the key innovations and environmental opportunities that allowed this adaptive radiation to take place. Return to Pianka lab page Page content... This page has hits |