|
The State of the Art in Community Ecology Eric R. Pianka ABSTRACT Many major advances have been made at various levels in biology, but the approach at the community level has stalled. Community ecology remains in its infancy and lags far behind other organismal and suborganismal disciplines due to its innate complexity, multi-dimensionality, extensive scale in space and time, multiple causality, as well as serious empirical difficulties of data collection and analysis. Pristine natural systems are vanishing rapidly before we understand how they are organized or how they behave. The study of communities must of necessity remain to some degree abstract and complex. Community ecology is extremely promising, important, and exceedingly urgent. Major new insights lie just around the corner. But community ecology is not for the faint at heart: it is one of the most challenging and most difficult of all sciences. Its vocabulary is still being developed. Identification of appropriate aggregate variables or macrodescriptors is essential, but constitutes a double-edged sword: macrodescriptors enable progress but simultaneously constrain its direction(s). At this early stage in community ecology, it is perhaps safest not to become overly "locked in" by words and concepts. A diversity of approaches seems desirable. Many community-level properties are probably simply epiphenomena that arise from pooling component populations: examples would presumably include trophic levels, guilds, subwebs, and ecological pyramids. Do communities also possess truly emergent properties that transcend those of mere collections of populations? For example, are patterns of resource utilization among coexisting species co-adjusted so that they mesh together in a meaningful way? If so, truly emergent community-level properties arise as a result of orderly interactions among component populations. This fundamental question needs to be answered. Either way, transcendent phenomena or epiphenomena cannot be studied at the level of individuals or populations. A major pitfall for community ecologists is that communities are not designed directly by natural selection (as are individual organisms). We must keep clearly in mind that natural selection operates by differential reproductive success of individual organisms. It is tempting, but dangerously misleading, to view ecosystems as "superorganisms" that have been "designed" for efficient and orderly function. However, antagonistic interactions at the level of individuals and populations (such as competition, predation, parasitism and even mutualisms) must frequently impair certain aspects of ecosystem performance while enhancing other properties. The traditional ecological approach to population interactions has been to consider species in pairs. While fruitful, this focus has diverted attention away from a more holistic systems-level approach. We must now make a determined effort to understand indirect as well as direct interactions within complex networks of interacting species. Properties of such networks themselves need to be evaluated. Approaches that incorporate mutualism and variable intensity in interactions must be developed. Competitive interactions should also be included and distinguished from the vertical ones that operate between trophic levels. Strong interactions may usually be more important than weak ones, but the cumulative effects of many weak interactions (as in diffuse competition and diffuse coevolution) could nevertheless be considerable. The extent to which indirect effects can and actually do balance direct effects needs to be ascertained. Whereas the reductionistic approach has been quite successful in other scientific disciplines, it will not lead to generalizations in community ecology. A holistic approach is required. A hybrid protocol for the analysis of community structure incorporates resource availability, electivities, nearness rank of neighbors in niche space, null models, and Monte Carlo statistical methodology. This new approach facilitates graphical comparisons of very different systems and is illustrated using assemblages of neotropical fish plus Australian and Kalahari desert lizards. Another neglected area with promise in community ecology is the effect of large-scale regional factors on the diversity and community structure at the local level scale. INTRODUCTION In biology, workers at different levels of approach look somewhat askance at the next higher level of approach, so that molecular biologists are slightly suspicious of cell biologists and cell biologists are just a little wary of physiologists. Physiologists are a bit suspicious of behaviorists and other whole-organism biologists, and it goes on. Unfortunately, many population ecologists distrust community ecology. The reason for this hesitancy to accept the next higher level may be that one must slur over interesting detail at one's own level in order to practice biology at the next level up. Interdisciplinary research and promotes tolerance and appreciation across the various levels of approach. Ecologists are not very interested in captive animals. Our subjects are wild organisms in natural settings, in some sort of a pristine natural situation, hopefully, with a normal environment to which that particular organism has become adapted and in which it has evolved. Rolston (1985) made a useful analogy, likening life on earth to a book written in a language that humans can barely read. Biologists are just now acquiring the skills necessary to read and decipher this book, but the poor book is tattered and torn, pages are missing (extinct species such as passenger pigeons), and entire chapters have been ripped out (e.g., the tall grass prairies of midwestern North America). There is a considerable urgency to study wild organisms in pristine natural habitats before they are gone forever. Molecular biologists can do experiments in test tubes or in small spaces in rooms in a matter of minutes or hours. Frequently it is possible to plan an experiment before lunch, execute it that afternoon and analyze the results that evening or the next day. As one ascends to higher and higher levels, larger areas are required and more time is needed. Satellite images are the electron micrographs of community ecologists. Community ecology requires thousands of square kilometers and decades or even centuries, making it quite an imposing subject. The scientific method has virtually become equated with the reductionistic approach. Scientists are trained first to break up whatever it is they are studying into its component parts and then to try to understand how those parts and components interact with one another. This very fruitful approach has led many important discoveries. For example, at the lowest level in biology are all the elegant biochemical findings: the discovery of DNA, the machinery of genetics, transcription, translation, etc. And as one ascends, equivalent discoveries have been made in cell biology and physiology. At the level of the whole organism, we have the concept of natural selection and important ideas about adaptive components of behavior. Even at the population level, a discovery of some importance has been made, namely the notion of kin selection. But at the community level, we remain abysmally ignorant. We cannot afford to allow community ecology to languish any longer. It has fallen well behind the other types of biology and needs to catch up. "Why has community ecology remained so primitive?" It is not for the impatient or faint at heart, and it requires a long time and a lot of space. Results do not come in overnight. Community ecology also demands considerable theoretical prowess because it is very abstract, conceptually difficult, and multi-dimensional. It is empirically intractable, too. A lot of devotion and hard work are necessary to collect any kind of data on an entire system or even a portion of one. Empirical problems of data collection and analysis are serious. Multiple causality may well be more of a problem in community ecology than it is at lower levels because its time- and space-scales are so vast. Many biologists, including population ecologists, find community ecology repugnant, and they shy away from it, for understandable reasons. Community ecology needs to attract population ecologists who are well versed in natural selection. It has become the province of systems ecologists and ecosystem engineers: population ecologists should become born-again community ecologists. Community ecology is doubtlessly one of the most difficult kinds of biology, but it has obvious utility as we approach oversaturation of this planet. Moreover, data must be gathered now because so many systems are vanishing. Community ecology is also very promising. Major new discoveries, potentially things as important as DNA and natural selection, remain undiscovered because biologists have shied away from this discipline. COMMUNITY ECOLOGY Community ecology is still struggling with its vocabulary. It remains a primitive and embryonic science. We are looking for macrodescriptors or aggregate variables that summarize some of the properties of systems; this is the usual reductionistic approach. Examples of macrodescriptors include relative abundance, species diversity, trophic levels, and food webs. Food webs have received considerable attention lately. Our choice of vocabulary constitutes a double-edged sword. Macrodescriptors selected to describe systems enable progress but simultaneously constrain directions. It may be best not to get too locked in until we are confident that we are going in the most fruitful directions. Moreover, a diversity of approaches seems desirable. Community ecology is full of pitfalls. A major trap is that it is tempting to view an ecosystem as a sort of a super organism, designed for smooth and efficient functioning. People make this mistake all the time. They lose sight of the fact that natural selection works at the level of individuals and that there are a multitude of antagonistic interactions among the members of a system, for example prey-predator interactions. Natural selection operating on predators may often reduce the stability of a system but should usually increase the efficiency of flow of energy up through the trophic levels, whereas selection operating on the prey has exactly the reverse effects. As a corollary, community-level properties of ecological efficiency and community stability may in fact be inversely related because natural selection operates at the level of the individual predators and prey. An interesting question in community ecology is "do communities really possess some kind of emergent properties that are more than mere statistical expressions of the component parts?" The latter sorts of phenomena have been termed "epiphenomena." Here's an example of a truly emergent property: If patterns of resource utilization among consumers in a system are co-adjusted with one another so that species A eats a lot of food type X, but species B eats only a little of X and a lot of food-type Y which in turn is not eaten by A, and this sort of complementary pattern permeates the whole system, then that system is organized and has some sort of structure. If a species could be removed, there would be a hole in the machine that would reflect what that species does (in fact, such an ecological vacuum might be quickly filled by shifts in patterns of resource utilization of the other species in the system). Like scientists in general, many community ecologists gravitate towards reductionism and they tend to approach communities from the bottom up. Communities can also be approached holistically from the top down. Tilman (1987) commented: "Must plant ecologists manipulate the densities of every species of plant, every soil bacterium, every fungus, every nematode, every herbivore, etc., and establish the effects of each manipulation on population densities and/or growth rates of all other species just to determine whether plants are actually competing with each other? Such an approach is so complex as to be ridiculous; yet that is the approach that a purely phenomenological perspective seems to require." Tilman threw up his hands at the complexity of entire natural communities. Pimm (1987) stated: "community ecology is becoming the ecology of guilds rather than the ecology of communities." Small pieces of a complex system may well lack important properties inherent in the intact system. In between the ecosystem systems approach and the guild ecology of microscopic community ecologists is community ecology of entire assemblages, subsets of the whole community, but larger than just guilds. Schoener (1986) argued that community ecology in the future will consist of a collection of theories, each with its power of prediction for a limited subset of species under specific environmental conditions. This kind of sentiment emerges from the microscopic perspective on communities. Diverse, complex systems cannot be neglected just because they're intractable. Some attempt must be made to deal with these phenomena holistically even though they are very messy. A great deal remains to be learned about community structure, even in the best studied systems. 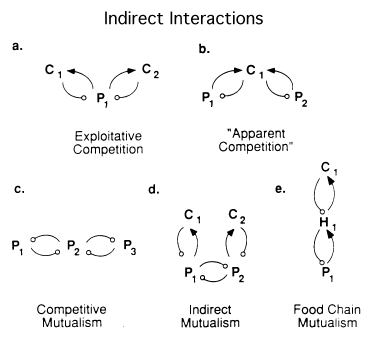 Still other kinds of mutualism can occur horizontally and vertically: competitive mutualism and food web or food chain mutualism. Ecologists have only recently begun to become aware of indirect interactions: for too long we have been overly simplistic and classified all interactions between species just into simple pairwise interactions. An entire multi-dimensional system, or a complex network, needs to be reduced a graphical state that elucidates the structure and organization of that entire system. "The mere attempt to define phenomena operationally can dramatically increase theory maturity." (Loehle 1987). Winemiller and I developed a hybrid approach to compare his aquatic systems with my own desert ones (Winemiller and Pianka, 1990). Our approach adopts a holistic perspective on a complex assemblage of interacting species and endeavors to represent the entire assemblage graphically to detect patterns of organization in that system. METHODOLOGY Resource utilization matrices constitute the raw data for analysis. A resource matrix contains a lot of vital information about a system. They identify which species eat which other species as well as which species are potential competitors because they share common foods. Such a matrix describes the food web structure of the system. Considerable tedious work is required to put together a satisfactory resource matrix. Statistical samples of all the species in the system must be collected; if the system is changing in time, this needs to be done quickly; to follow changes in the community through time, adequate samples at different times are necessary. Entries in the resource matrix are used to calculate probabilities. These vary between zero and one, reflecting the probability that a given consumer species, say 1, will use a particular resource state, say 2. Some utilization probabilities, or uij's, in the matrix will obviously be zeros because some consumers don't use some resources. Simple dietary proportions, or pi's, weight uncommon or very abundant resources disproportionately. Resource utilization can be standardized in terms of relative availabilities (Ivlev 1961) . Resource availabilities are not easily measured in the field. Insects can be sampled with sweep nets, DeVac vacuum cleaners, tanglefoot sticky traps, pit traps, or burliese funnels: each technique yields very different results. Some insects are simply more easily pit trapped than others, whereas others are captured by burliese funnels more than others, etc. Pefaur and Duellman (1980; pers. comm.) studied Andean herps from Columbia to Argentina. They fenced study plots and collected all frogs and squamates (lizards and snakes); inside these plots, all conspicuous insects that a human observer could find were also collected, and saved with the intention of using these as standards against the stomach contents of the herps. Humans collected very few of the insects eaten by the herpetofaunas, only about 10% (Duellman, pers. comm.). Incredibly enough, 90% of the insects that were in stomachs in fact were not even collected by diligent humans! It is a gross and dangerously misleading oversimplification to accept the idea that a single resource vector exists out there that describes a system. Each species experiences its own resource availabilities that depend to some extent on that species' use of space and time as well as its behavior and foraging mode. Various solutions to this problem have been proposed. One technique weights resources in proportion to their use in the overall system. Resource totals in the resource matrix can be considered a sort of bioassay of resource availability. In a system of a hundred species, diets summed over all component species represent an estimated resource availability vector. Probabilistic analogues of electivity are computed and an analysis can proceed that is unbiased by resource availability (for greater detail, see Winemiller and Pianka, 1990). A classic paper on community ecology and resource partitioning among Thailand herps was published in 1977 (Inger and Colwell 1977). These authors pointed out that no consensus exists as to how to approach community ecology, and they said there is "no standard protocol for community ecology." Those statements are still true today, a dozen years later. Inger and Colwell (1977) suggested a nearest neighbor approach to look at communities; each species' overlap with every other species is ranked from the closest neighbor in niche space to those increasingly more distant. These generate monotonically declining curves for all the species in the system. Some species have high overlap well out into niche space whereas overlap in others falls off rapidly (such consumers are very distinct with low overlap with most of the other members of the system). We used simply the mean overlap at a given rank across all species in the system. A system in the Kalahari desert with 15 species of lizards can be represented with a single curve, simply the arithmetic average over all fifteen species at each rank in niche space. NULL MODELS Another promising technique involves null models (Colwell and Winkler, 1984). One of the greatest challenges is to find something with which to compare a given community. It is extremely difficult to compare a system with someone else's system: that is in fact what provoked us to devise these techniques, to compare the fish with the lizards. Elements of a resource matrix can be scrambled according to some rules (Sale 1974) to create "pseudo-communities"; these are then compared with the original prototype to look for differences to see how the original system is in fact organized. Sale's algorithm involved scrambling all the utilization coefficients, whatever their values are for each consumer in the system (zeros or positive). All its elements for each consumer are randomly rearranged. Rearranged utilization probability u31 could fall with equal probability into any slot in that species' utilization vector and u11 would do the same. With a computer, one can easily perform this rearrangement a hundred times, and use a bootstrap approach and Monte Carlo statistics to generate a distribution against which the observed can be compared. Statistics can be computed to discern whether or not any differences are significant. A slightly different algorithm which turns out to be equally instructive was suggested by Lawlor (1980b): this algorithm leaves the zero structure of the resource matrix intact. So that if consumer 1 does not eat resource state 3, a zero must remain in cell u31; it is frozen and not allowed to change. Elements in the resource matrix are scrambled but only among the resources that are actually used by a given species. (Lawlor's method is termed the "conserved-zero" approach and Sale's method the "scrambled-zero" algorithm because it destroys the zero structure.) BENCH TESTS To exploit these techniques on real fish and lizard systems (Winemiller and Pianka, 1990), we constructed a test set of hypothetical model systems that had understandable known structure. Model systems both with and without guilds were "bench tested" with our methodology. Three systems with two guilds of equal size, five species in each, were constructed. We examined how the randomization algorithms affected these simple model systems. At the top are two guilds with very high, almost total, overlap. At the bottom are two guilds with low overlap. In the middle, overlap is intermediate. When zeros are scrambled, of course, guild structure is destroyed and the result is that the scrambled-zero algorithm results in increased overlap at distant ranks in the niche space. Effectively they float, because the original system (the prototype) had niche segregation in it which was destroyed when the resource matrix elements were scrambled. We assembled another set of three systems with guilds of different sizes; these behave somewhat the same. We also put together systems like these without any guild structure, but with resource partitioning. It became harder to get pseudo-communities to float, although some conserved zero pseudo-communities did float, which we interpret as evidence of niche segregation. We were also interested in the phenomenon of core resources. Both fish and lizards exploit certain core resources extensively. Among lizards, these are termites and ants, especially termites. Among fish, mayflies constitute a core resource. We created some systems with extensive or total overlap on certain core resources and then unique resources used by each species that were partitioned. Effects of these algorithms can be summed up: With consumers piled up on a certain resource state (core resources or guilds with everything within a guild eating the same things), the scrambled-zero algorithms tended to sink, and fall below the observed. But, when resources are partitioned, the conserved-zero pseudo-communities tend to be above the observed system (i.e., they "float"). RESULTS FROM REAL SYSTEMS Winemiller studied ichthyofaunas of aquatic systems in Venezuela and in Costa Rica. One of his study sites has over eighty species of fish in it over the course of an entire annual cycle. Winemiller discovered how to collect virtually an entire freshwater aquatic assemblage. We pulled a seine through Ca�o Maraca during the dry season and captured over a thousand fish of dozens of species (plus a "bonus" of a couple of large ca�man!) His sample sizes are on the order of three hundred to five hundred. Winemiller went through statistical subsamples and separated his data into wet versus dry season resource matrices. Prey content by volume was estimated to the finest discriminatory abilities possible, given our own expertise, usually insect orders. We actually examined 18 different resource matrices, with two or three from each of eight sites: a wet and dry season for each of four fish sites and microhabitat plus diet matrices for each of four lizard sites. Numbers of fish species on these sites vary from 19 to 59 and numbers of lizard species vary from 15 to 39. We had between 40 and 217 resource states among the sites analyzed. An Australian desert site, the L-area near Laverton, Western Australia, has 32 species of lizards. My favorite study area is Red Sands, near Yamarna Homestead in Western Australia. I have collected 46 species of lizards there so far and expect to find several more. The hummock grass tussock plant growth form (spinifex) is very important in the Australian desert. These tussocks, as large as a meter in diameter, house certain lizards that virtually never leave them. Some lizards are highly adapted to spinifex and swim through it with ease. Each lizard collected, some 3000 in Australia and another 2000 from the Kalahari, was weighed and measured in the field, individually tagged, and then permanently preserved by injection with formaldehyde. These specimens are all safely deposited in major museums where they are valuable material for systematics research. Many of my lizards have been dissected by people interested in functional anatomy. When the lizards are returned eventually to the laboratory, each lizard is measured--ten different body measurements are made on them for anatomical analyses, and then each lizard is dissected and its reproductive state is noted, and relative clutch mass is estimated (testicular cycles can be deduced from serial samples like this), but the most important thing for present purposes is that stomachs are pulled. A competent entomologist went through the stomach contents on the Australian lizards, identifying 100,000 or more prey items to the finest categories possible. Costa Rican fish assemblages are shown in Figure 11 during two seasons, wet and dry, based on two different resource matrices (some fish species present in the wet are not there during the dry season). Mean overlap in the observed system is shown in the upper panel of each graph with solid circles. Overlap at each rank in niche space is plotted, the average similarity between consumers at the first, second, third, etc. rank. Pseudo-communities are shown with the open symbols, the conserved zeros are open circles; the scrambled zeros are open triangles. In the lower panels of each figure, the percentage of pseudo-communities that either float or sink are plotted. In this case, sinking of the scrambled zero pseudo-communities is interesting, as is floating of conserved zero pseudo-communities which reflects niche segregation. The dashed lines in the bottom panels are at 5% and 95% confidence levels, so when a pseudo-community lies above the upper dashed line or below the lower dashed line, there is a statistically significant difference between the pseudo-communities and the observed system. At close ranks in niche space, conserved-zero systems don't float but farther out they clearly differ from observed systems and to some extent, except for the lower right panel in the figure, the scrambled-zero psuedo-communities almost invariably sink, at most ranks. Venezuelan fish float and sink even better (Fig 12). Realize that these represent an enormous amount of information in one graph. In one case, at Maraca (top panel), 29,000 fish went into the figure. Winemiller lived down there for a full year, collected many thousand fish, brought them back and spent an entire year going through vast numbers of stomachs. All this information can be represented on a single page with a simple graph that one can examine and interpret with a little bit of training. These aquatic systems are highly organized, with guild structure, core resources, plus niche segregation. Consumers are piled up on certain core resources, reflected in the sinking of the scrambled-zero pseudo-communities. But those same consumers are also segregated out on those core resources that they do use, with different species using the same core resources but with different probabilities. Australian lizards are depicted in Figure 13. At the very top are microhabitats, in the middle are standard dietary resource matrices (nineteen prey categories, largely insect orders). At the bottom are expanded dietary resource matrices with 201 different prey categories represented at Red Sands and 217 prey categories recognized at the L area on the right. Some interesting things emerge from these plots. Scrambled-zero pseudo-communities tend to sink in all cases which indicates piling up on certain core resources and is indicative of guild structure. Conserved-zero pseudo-communities float pretty well in microhabitats which indicates niche segregation: different species use different microhabitats and they float fairly unequivocally, except at the closest ranks at Laverton. In the middle plot, though, pseudocommunities do not show very much floating because food resource states are too crudely differentiated resulting in a piling up of consumers on some resource states. The same data are shown in the bottom panel, but with finer discrimination of prey resource states: note that conserved zero pseudocommunities float as they did in the fish, indicating segregation. Kalahari desert systems are more loosely organized than those in the Australian desert. There are fewer lizard species in the Kalahari and prey could not be distinguished to categories as fine as those in Australia: only 46 different prey resource states were recognized. For microhabitats, there is some hint of floating in the conserved zero pseudo-communities at Tsabong, Botswana, but not at Bloukrans, South Africa. Examining diet shows there is not much niche segregation. There isn't any at all at Tsabong, but there seems to be some at more distant ranks, at Bloukrans. All in all, the systems we examined tend to be fairly highly organized. This technique should facilitate analyses of other systems and allow comparisons with our own. CONCLUSIONS Community ecology has for too long been perceived as repugnant and intractably complex by reductionistic population ecologists. As a result, the discipline has been neglected and now lags far behind the rest of biology. As we approach oversaturation of this planet, we will soon be needing all the ecological understanding and wisdom that we can possibly muster. No longer can we afford to remain ignorant of the principles and factors affecting the structure and function of entire systems of interacting species. Community ecology is not for the faint at heart, but its serious students may well be rewarded with major new advances and discoveries as exciting and important as DNA and natural selection. I'd like to end with a plea and ask all of you to be tolerant of community ecology. Allow the discipline to try to catch up with the rest of biology which has enjoyed such a head start. I would particularly like to ask those community ecologists who adopt the microscopic approach, that is, "experimental" community ecologists, to be tolerant of community ecologists trying to study entire assemblages and who are adopting a more holistic and "descriptive" approach to communities. We must seek and maintain dialogue and mutual respect as we work towards integrating our divergent but complementary approaches. |