
Indirect Interactions Between Species
One of the most interesting questions in community ecology is the question of whether or not communities really possess some kind of emergent properties that are more than mere statistical expressions of the component parts. The latter sorts of phenomena have been termed "epiphenomena." Here's an example of a truly emergent property. If patterns of resource utilization among consumers in a system are co-adjusted with one another so that species A eats a lot of food type X, but species B eats only a little of X and a lot of food-type Y which in turn is not eaten by A, and this sort of complementary pattern permeates the whole system, then that system is organized and has some sort of structure. If we could remove a species, there would be a hole in the machine that would reflect what that species does (in fact, such an ecological vacuum might be quickly filled by shifts in patterns of resource utilization of the other species in the system).

Like scientists in general, many community ecologists gravitate towards reductionism and they tend to approach communities from the bottom up. I would like to make a case for approaching communities holistically from the top down. We need the microscopic experimental approach but we should also examine other approaches. A plant ecologist, Tilman (1987) recently made this statement: "Must plant ecologists manipulate the densities of every species of plant, every soil bacterium, every fungus, every nematode, every herbivore, etc., and establish the effects of each manipulation on population densities and/or growth rates of all other species just to determine whether plants are actually competing with each other? Such an approach is so complex as to be ridiculous; yet that is the approach that a purely phenomenological perspective seems to require." Tilman threw up his hands at the complexity of entire natural communities. Many reductionistic microscopic community ecologists have done just that. In the anuran community ecology symposium at this conference, we heard about the biology of small groups or sets of species. An example would be the work of Hairston (1987) on salamander guilds. In a book review entitled "Putting the species back into community ecology," Pimm (1987) put it in this way: "community ecology is becoming the ecology of guilds rather than the ecology of communities." Small pieces of a complex system may well lack important properties inherent in the intact system. I propose to consider something in between the ecosystem systems approach and the guild ecology of microscopic community ecologists and that is community ecology of entire assemblages, subsets of the whole community, but larger than just guilds.

that has converged on the viperid body plan.
Recently Schoener (1986) argued that community ecology in the future will consist of a collection of theories, each with its power of prediction for a limited subset of species under specific environmental conditions. This kind of sentiment emerges from the microscopic perspective on communities. In my opinion, we cannot afford to neglect diverse, complex systems just because they're intractable. We must make some attempt to deal with these phenomena holistically even though they are very, very messy. A great deal remains to be learned about community structure, even in the best studied systems (Menge and Farrell, 1989).
In complex networks of interacting species, numbers of indirect interactions (Kerfoot and Sih, 1987), that is, interactions that require paths of length more than one, goes up exponentially with species diversity (Patten, 1983). Of course direct interactions potentially increase faster and faster with more species too. Whereas direct interactions are readily understood, indirect interactions can lead to counterintuitive situations that have been called, for example, "apparent competition" (Fig. 5b; Holt, 1977). When a single predator has two prey species, if a removal experiment is performed on one of the prey, the predator must increase its predation on the other species. A naive community ecologist might interpret the response as competition when it isn't competition at all! Other such indirect interactions involve more than just two links (Fig. 5). One I find most interesting is in the bottom center (Fig. 5d), where two consumers are not competing or interacting directly in any way, but because their prey are competing at the next trophic level down, a positive apparent mutualism emerges between the two consumers that has been called facilitation or indirect mutualism (Vandermeer, 1980).
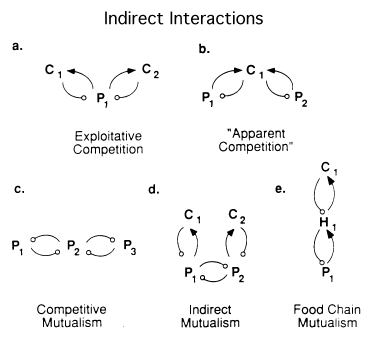
Indirect interactions in communities. (a) Two consumers sharing a common prey may compete indirectly via classical exploitation competition (resource depression). (b) Two prey species may only appear to compete, because if either increases, a shared predator also increases, which operates to the detriment of the other prey population (Holt 1977). (c) Populations of three species at the same trophic level, arranged so that one species, P(2) is sandwiched between two others. Populations P(1) and P(3) are indirect mutualists because each inhibits the other's competitor -- P(2). (d) A more complex four-species system that results in an indirect "mutualism" or "facilitation" (Vandermeer 1980). In this case, populations C(1) and C(2), which do not interact directly, but consume different prey species, interact indirectly because their prey compete: if consumer C(1) increases, its prey P(1) decreases, which in turn reduces competition with P(2), providing more food for consumer species C(2). (e) Populations of three species at three trophic levels: the plant P(1) and the carnivore C(1) are indirect mutualists because they share an herbivore H(1). Numerous other sorts of indirect interactions are also possible.
Still other kinds of mutualism can occur horizontally and vertically: competitive mutualism (Pianka, 1981: Fig. 5c) and food web or food chain mutualism (Fig. 5e). Ecologists have only recently begun to become aware of indirect interactions: for too long we have been overly simplistic and classified all interactions between species just into simple pairwise interactions (Bender et al., 1984; Brown et al., 1986; Pianka, 1987).
We need ways of taking an entire multi-dimensional system, or a complex network, and reducing it to some kind of a graphical state in which we can begin to appreciate the structure and organization of that entire system. Loehle (1987) said "the mere attempt to define phenomena operationally can dramatically increase theory maturity."
(Back to Guild Structure and Community Organization, click here)