MICROBIAL
MODELS:
THE GENETICS
OF VIRUSES AND BACTERIA
Viruses and bacteria are the
simplest biological systems serve as microbial models.
- Microbiologists provided
most of the evidence that genes are made of DNA, and they worked
out most of the major steps in DNA replication, transcription,
and translation.
Bacteria are prokaryotic organisms, with cells much smaller and
more simply organized. Fig
18.1.
Viruses are smaller and simpler still, being little more than
genes in a protein coat.
Viral genetics
- Researchers discovered viruses
by studying a plant disease
Tobacco
mosaic disease stunts the growth and mottles plant leaves.
Several scientists determined that the disease could be transmitted
from plant to plant by spraying the sap from an infected plant
to a healthy one, even after the sap had been filtered through
a filter that should have removed bacteria. It was also determined
that it could reproduce only within the host, could not be cultivated
on nutrient media, and was not inactivated by alcohol, generally
lethal to bacteria.
In 1935, Stanley crystallized the pathogen, the tobacco
mosaic virus (TMV). Fig
18.9b.
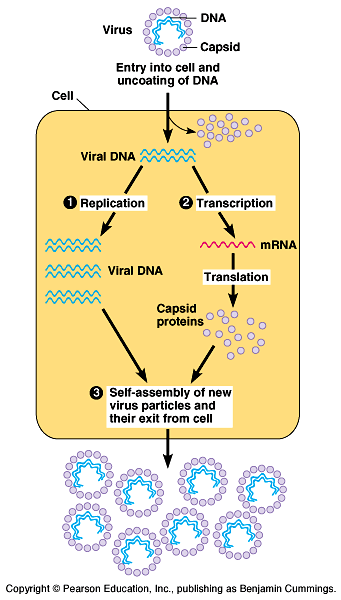
A virus is a genome
enclosed in a protective coat
- Viruses are not cells. They
are infectious particles consisting of nucleic acid encased in
a protein coat, and, in some cases, a membranous envelope. Fig.
18.2.
Viral genomes may consist of double-stranded DNA, single-stranded
DNA, double-stranded RNA, or single-stranded RNA, depending on
the specific type of virus. The smallest viruses have only four
genes, while the largest have several hundred.
The capsid is a protein shell enclosing the viral genome.
Capsids are built of a large number of protein subunits called
capsomeres, but with limited diversity.
- Some viruses have viral
envelopes, membranes cloaking their capsids.
These envelopes are derived from the membrane of the host cell.
They also have some viral proteins and glycoproteins.
The most complex capsids are found in viruses that infect bacteria,
called bacteriophages
or phages. T-even
phages may infect E. coli.
Viruses can reproduce
only within a host cell.
Fig
18.3.
- Viruses are obligate intracellular
parasites. Viruses lack the enzymes for metabolism or ribosomes
for protein synthesis.
Each type of virus can infect and parasitize only a limited range
of host cells, called its host range.
Viruses identify host cells by a "lock-and-key" fit
between proteins on the outside of virus and specific receptor
molecules on the host's surface.
Some viruses (like the rabies virus) have a broad enough host
range to infect several species, while others infect only a single
species.
Most viruses of eukaryotes attack specific tissues.
Human cold viruses - upper respiratory tract.
The AIDS virus - white blood cells.
A viral infection begins when the genome of the virus enters
the host cell.
Once inside, the viral genome commandeers its host, reprogramming
the cell to copy viral nucleic acid and manufacture proteins
from the viral genome.
The nucleic acid molecules and capsomeres then self-assemble
into viral particles and exit the cell.
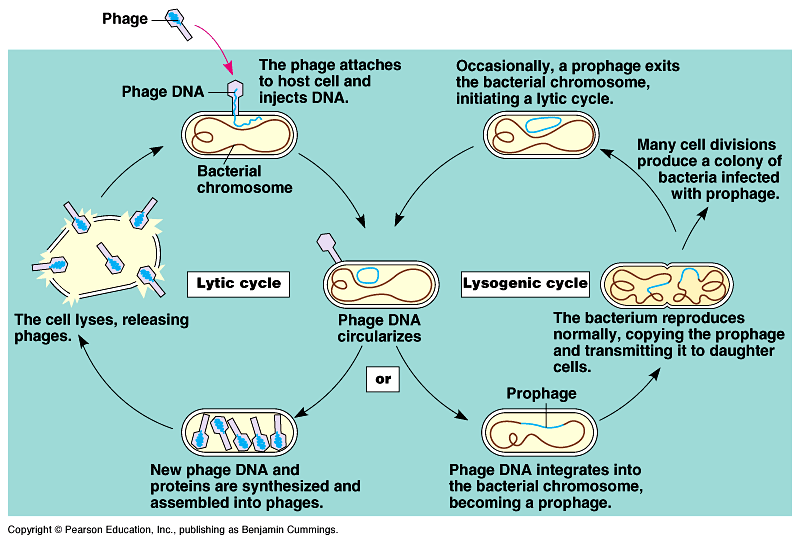
Phages reproduce using
lytic or lysogenic cycles
- Some double-stranded DNA
viruses can reproduce by two alternative mechanisms: the lytic
cycle and the lysogenic cycle.
In the lytic cycle (Fig.
18.4), the phage reproductive cycle culminates in the death
of the host, when the bacterium lyses and releases the phages
produced to infect others.
-
- Virulent phages reproduce only by a lytic cycle.
Bacterial defenses:
- 1. Natural selection favors
bacterial mutants with receptors sites that are no longer recognized
by a particular type of phage.
2. Bacteria produce restriction nucleases that recognize
and cut up foreign DNA, including certain phage DNA.
3. Modifications to the bacteria's own DNA prevent its destruction
by restriction nucleases.
In the lysogenic cycle, the phage genome replicates without
destroying the host cell.
Temperate phages, like phage lambda, use both lytic
and lysogenic cycles. Fig
18.5. Movie!
- During a lytic cycle, the
viral genes immediately turn the host cell into a virus-producing
factory, and the cell soon lyses and releases its viral products.
During the lysogenic cycle, the viral DNA molecule, is incorporated
by genetic recombination into a specific site on the host cell's
chromosome.
In this prophage stage, one of its genes codes for a protein
that represses most other prophage genes.
Every time the host divides, it also copies the viral DNA and
passes the copies to daughter cells.
Occasionally, the viral genome exits the bacterial chromosome
and initiates a lytic cycle.
This switch from lysogenic to lytic may be initiated by an environmental
trigger.
Animal viruses very
diverse
One key variable is the type of nucleic acid that serves
as a virus's genetic material.
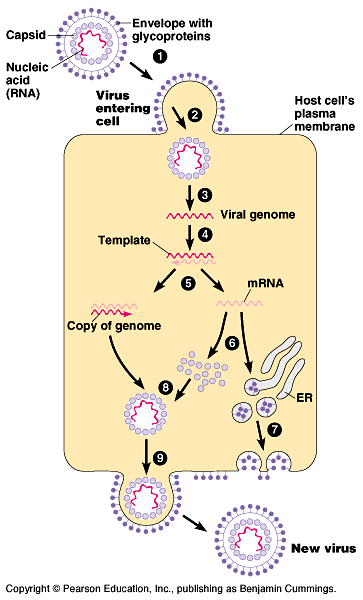
Another variable is the presence or absence of a membranous
envelope. Viruses equipped with an outer envelope use the
envelope to enter the host cell. Fig
18.6.
Glycoproteins on the envelope bind to specific receptors
on the host's membrane. The envelope fuses with the host's membrane,
transporting the capsid and viral genome inside.
The viral genome duplicates and directs the host's protein synthesis
machinery to synthesize capsomeres with free ribosomes
and glycoproteins with bound ribosomes.
After the capsid and viral genome self-assemble, they bud
from the host cell covered with an envelope derived from the host's
plasma membrane, including viral glycoproteins. These enveloped
viruses do not necessarily kill the host cell.
Some viruses have envelopes that are not derived from plasma membrane.
The envelope of the herpesvirus is derived from the nuclear
envelope of the host.
These double-stranded DNA viruses reproduce within the
cell nucleus using viral and cellular enzymes to replicate and
transcribe their DNA.
Herpesvirus DNA may become integrated into the cell's genome as
a provirus, which remains latent within the nucleus
until triggered by physical or emotional stress to leave the genome
and initiate active viral production.
In some with single-stranded RNA (class IV), the genome
acts as mRNA and is translated directly.
In others (class V), the RNA genome serves as a template
for mRNA and for a complementary RNA. This complementary strand
is the template for the synthesis of additional copies of genome
RNA.
All viruses that require RNA -> RNA synthesis to make mRNA
use a viral enzyme that is packaged with the genome inside the
capsid.
Retroviruses (class VI) have the most complicated
life cycles.
- These carry an enzyme, reverse
transcriptase, which transcribes DNA from an RNA template.
The newly made DNA is inserted as a provirus into a chromosome
in the animal cell.
The host's RNA polymerase transcribes the viral DNA into more
RNA molecules.
These can function both as mRNA for the synthesis of viral proteins
and as genomes for new virus particles released from the cell.
Human immunodeficiency virus (HIV), the virus that
causes AIDS (acquired immunodeficiency syndrome)
is a retrovirus. Fig.
18.7.
The viral particle includes an envelope with glycoproteins
for binding to specific types of white blood cells, a capsid
containing two identical RNA strands as its genome and
two copies of reverse transcriptase.
Some viruses damage or kill
cells by triggering the release of hydrolytic enzymes from
lysosomes.
Some viruses cause the infected cell to produce toxins
that lead to disease symptoms.
Others have molecular components, such as envelope proteins,
that are toxic.
In some cases, viral damage is easily repaired (respiratory epithelium
after a cold), but in others, infection causes permanent damage
(nerve cells after polio).
Many of the temporary symptoms associated with a viral infection
results from the body's own efforts at defending itself against
infection.
Vaccines.
- The first vaccine, using
cowpox, was developed in the late 1700s by Edward Jenner
to prevent smallpox.
Many others have since been developed.
Antibiotics don't work against viruses.
Antivirals - recently developed drugs to combat some viruses,
mostly by interfering with viral nucleic acid synthesis.
AZT interferes with reverse transcriptase of HIV.
Acyclovir inhibits herpesvirus DNA synthesis.
Emergent viruses. Fig
18.8.
- HIV, the AIDS virus, seemed
to appear suddenly in the early 1980s.
The deadly Ebola virus has caused hemorrhagic fevers in central
Africa periodically since 1976.
The emergence of these new viral diseases is due to three
processes:
mutation
RNA viruses have high mutation rates because replication of their
nucleic acid lacks proofreading.
Influenza strains
spread of existing viruses from one species to another
It is estimated that about three-quarters of new human diseases
have originated in other animals.
For example, hantavirus, which killed dozens of people
in 1993, normally infects rodents, especially deer mice.
Dissemination of a viral disease from a small, isolated population.
AIDS, present only in small populations in Africa, went unnamed
and unnoticed for decades before spreading around the world.
Affordable international travel, blood transfusion technology,
sexual promiscuity, and the abuse of intravenous drugs, allowed
a previously rare disease to become a global scourge.
Tumor viruses include
retrovirus, papilloma virus, adenovirus, and herpesvirus types.
- The hepatitis B virus is
associated with liver cancer.
The Epstein-Barr virus, which causes infectious mononucleosis,
has been linked to several types of cancer in parts of Africa,
notably Burkitt's lymphoma.
Papilloma viruses are associated with cervical cancers.
The HTLV-1 retrovirus causes a type of adult leukemia.
All tumor viruses transform cells into cancer cells after integration
of viral nucleic acid into host DNA.
Viruses may carry oncogenes that trigger cancerous characteristics
in cells.
These oncogenes are often versions of proto-oncogenes
that generally code for growth factors or proteins involved in
growth factor function.
In other cases, a tumor virus transforms a cell by turning on
or increasing the expression of proto-oncogenes.
It is likely that most tumor viruses cause cancer only in combination
with other mutagenic events.
Plant viruses Fig
18.9.
- Most are RNA viruses with
rod-shaped capsids produced by a spiral of capsomeres.
Plant viral diseases are spread by two major routes.
In horizontal transmission, a plant is infected with the
virus by an external source.
In vertical transmission, a plant inherits a viral infection
from a parent.
This may occurs by asexual propagation or in sexual reproduction
via infected seeds.
Viroids and prions
- Viroids, smaller and simpler than even viruses,
consist of tiny molecules of naked circular RNA that infect plants.
Their several hundred nucleotides do not encode for proteins
but can be replicated by the host's cellular enzymes.
Prions are infectious proteins that spread a disease.
They are thought to cause several degenerative brain diseases
including scrapie in sheep, "mad cow disease," and
Creutzfeldt-Jacob disease in humans.
According to the leading hypothesis, a prion is a misfolded form
of a normal brain protein, which then converts normal proteins
into the prion version. Fig
18.10.
Viruses living or nonliving?
- An isolated virus is biologically
inert and yet it has a genetic program written in the universal
language of life.
Because viruses depend on cells for their own propagation, they
evolved after the first cells appeared, probably fragments
of cellular nucleic acids that could move from one cell to another.
A viral genome usually has more in common with the genome of
its host than with those of viruses infecting other hosts.
The evolution of capsid genes may have facilitated the infection
of undamaged cells.
Candidates for the original sources of viral genomes include
plasmids, small, circular DNA molecules that are separate
from chromosomes, and transposons, DNA segments that can
move from one location to another within a cell's genome. Both
are mobile genetic elements.
The Genetics of Bacteria
- Bacteria are very adaptable,
both in the evolutionary sense of adaptation via natural selection
and the physiological sense of adjustment to changes in the environment
by individual bacteria.
The major component of the bacterial genome is one double-stranded,
circular DNA molecule.
Tight coiling of the DNA results in a dense region of DNA, called
the nucleoid, not bound by a membrane.
Many bacteria have plasmids, much smaller circles of DNA.
Bacterial cells divide by binary fission. Fig
18.11.
New mutations, though individually rare, can have a significant
impact on genetic diversity when reproductive rates are very
high because of short generation spans.
Genetic recombination
- In bacteria, the combining
of DNA from two individuals into a single genome.
Recombination occurs through three processes:
Transformation
Transduction
Conjugation
- Transformation is the alteration of a bacterial
cell's genotype by the uptake of naked, foreign DNA from the
surrounding environment.
For example, harmless Streptococcus pneumoniae bacteria
can be transformed to pneumonia-causing cells, when a live nonpathogenic
cell takes up a piece of DNA that happens to include the allele
for pathogenicity from dead, broken-open pathogenic cells. Fig
18.12.
Many bacterial species have surface proteins that are specialized
for the uptake of naked DNA.
Transduction occurs when a phage carries bacterial genes
from one host cell to another. Fig
18.13.
In generalized transduction, a small piece of the host
cell's degraded DNA is packaged within a capsid, rather than
the phage genome.
When this phage attaches to another bacterium, it will inject
this foreign DNA into its new host.
This type of transduction transfers bacterial genes at random.
Specialized transduction occurs via a temperate phage.
When the prophage viral genome is excised from the chromosome,
it sometimes takes with it a small region of adjacent bacterial
DNA.
These bacterial genes are injected along with the phage's genome
into the next host cell.
Specialized transduction only transfers those genes near the
prophage site on the bacterial chromosome.
Both generalized and specialized transduction use phage as a
vector to transfer genes between bacteria.
Conjugation transfers genetic material between two bacterial
cells that are temporarily joined. Fig
18.15.
One cell ("male") donates DNA and its "mate"
("female") receives the genes.
A sex pilus (Fig
18.14) from the male initially joins the two cells and creates
a cytoplasmic bridge between cells.
"Maleness," the ability to form a sex pilus and donate
DNA, results from an F factor as a section of the bacterial
chromosome or as a plasmid.
Plasmids, including the F plasmid,
are small, circular, self-replicating DNA molecules.
Episomes, are a genetic element that can exist as a plasmid
or as part of a chromosome. Episomes, like the F plasmid, can
undergo reversible incorporation into the cell's chromosome.
Temperate viruses also qualify as episomes.
Plasmid genes are advantageous in stressful conditions.
Recombination exchanges segments of DNA.
This recombinant bacteria has genes from two different cells.
Antibiotic resistance.
- The genes conferring resistance
are carried by plasmids, specifically the R plasmid (R
for resistance).
Some of these genes code for enzymes that specifically destroy
certain antibiotics, like tetracycline or ampicillin.
When a bacterial population is exposed to an antibiotic, individuals
with the R plasmid will survive and increase in the overall population.
Because R plasmids also have genes that encode for sex pili,
they can be transferred from one cell to another by conjugation.
A transposon is a piece
of DNA that can move from one location to another in a cell's
genome.
- Transposon movement occurs
as a type of recombination between the transposon and another
DNA site, a target site.
In bacteria, the target site may be within the chromosome, from
a plasmid to chromosome (or vice versa), or between plasmids.
Transposons can bring multiple copies for antibiotic resistance
into a single R plasmid by moving genes to that location from
different plasmids.
This explains why some R plasmids convey resistance to many antibiotics.
Some transposons (so called "jumping genes")
do jump from one location to another (cut-and-paste translocation).
The simplest bacterial transposon, an insertion sequence,
consists only of the DNA necessary for the act of transposition.
Fig
18.16.
The insertion sequence consists of the transposase gene,
flanked by a pair of inverted repeat sequences.
The transposase enzyme recognizes the inverted repeats
as the edges of the transposon and cuts the transposon from its
initial site and inserts it into the target site. Fig
18.17.
Insertion sequences cause mutations when they happen to
land within the coding sequence of a gene or within a DNA region
that regulates gene expression.
Composite transposons (complex transposons) include extra
genes sandwiched between two insertion sequences. Fig
18.18.
Composite transposons may help bacteria adapt to new environments.
Repeated movements of resistance genes by composite transposition
may concentrate several genes for antibiotic resistance onto
a single R plasmid.
Transposable genetic elements are important components of eukaryotic
genomes as well.
In the 1940s and 1950s Barbara McClintock investigated changes
in the color of corn kernels, proposing that the changes in kernel
color only made sense if mobile genetic elements moved from other
locations in the genome to the genes for kernel color.
When these "controlling elements" were inserted next
to the genes for kernel color, they would activate or inactivate
those genes.
Individual bacteria
may adjust their metabolism to cope with environmental change
Fig
18.19
- They may vary the number
of specific enzyme molecules
by regulating gene expression.
They may adjust the activity of enzymes already present.
The operon model for
the control of gene expression in bacteria.
- An operon consists
of three elements:
The genes that it controls.
A promotor region where RNA polymerase first binds.
An operator region between the promotor and the first
gene that acts as an "on-off switch."
Repressible enzymes generally function in anabolic
pathways, synthesizing end products.
When the end product is present in sufficient quantities, the
cell can allocate its resources to other uses.
- The trp operon is
an example of a repressible operon, one that is inhibited
when some tryptophan molecules bind as corepressors
to the repressor protein.
Tryptophan absent, repressors inactive, gene transcribed transcribed.
Fig
18.20a
Tryptophan present, repressor
active, gene not transcribed. Fig
18.20b.
An inducible operon
is stimulated when a specific small molecule interacts
with a regulatory protein.
Inducible enzymes usually function in catabolic pathways,
digesting nutrients to simpler molecules.
By producing the appropriate enzymes only when the nutrient is
available, the cell avoids making proteins that have nothing to
do.
- The lac operon
contains genes that code for enzymes that digest lactose.
In the absence of lactose, this operon is off as an active repressor
binds to the operator and prevents transcription. Fig
18.21a.
When lactose is present
in the cell, the repressor is inactive and the operon is on.
Fig
18.21b.
Both repressible and inducible operons demonstrate negative
control because active repressors can only have negative
effects on transcription.
Positive gene control occurs when an activator molecule
interacts directly with the genome to switch transcription
on.
- Even if the lac operon
is turned on, the degree of transcription depends on the concentrations
of other substrates. Cellular metabolism first uses glucose.
If glucose levels are low (along with overall energy levels),
then cyclic AMP (cAMP) binds to cAMP receptor
protein (CRP) which activates transcription. Fig
18.22a
If glucose levels are sufficient and cAMP levels are low (lots
of ATP), then the CRP protein has an inactive shape and
cannot bind upstream of the lac promotor. Fig
18.22b
The lac operon will be transcribed but at a low level.
For the lac operon, the presence / absence of lactose
determines if the operon is on or off.
Overall energy levels in the cell determine the level of transcription,
a "volume" control, through CRP.
CRP works on several operons that encode enzymes used in catabolic
pathways.
If glucose is present and CRP is inactive, then the synthesis
of enzymes that catabolize other compounds is slowed.
If glucose levels are low and CRP is active, then the genes which
produce enzymes that catabolize whichever other fuel is present
will be transcribed at high levels.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}