|
Fire Succession on the B-area 
© Eric R. Pianka  The very flammable hummock grass plant life form is unique to Australia.
Spinifex tussocks are almost perfectly designed for combustion, consisting
of hemispherical clumps of numerous match stick sized, curled blades of dry
grass filled with flammable resins, loosely interpenetrating one another and
laced with ample air spaces. With such ideal tinder (on a scale of 1 to 10
as as measure of perfect tinder, spinifex gets a 9), one match will often
start a raging bonfire. Spinifex has been called an ideal pyrophyte.
Spinifex is an exceedingly important habitat element for many species of
Australian desert lizards. Individuals of most species take refuge in spinifex
from predators and the sun. Some species occur only in large long unburned tussocks.
The very flammable hummock grass plant life form is unique to Australia.
Spinifex tussocks are almost perfectly designed for combustion, consisting
of hemispherical clumps of numerous match stick sized, curled blades of dry
grass filled with flammable resins, loosely interpenetrating one another and
laced with ample air spaces. With such ideal tinder (on a scale of 1 to 10
as as measure of perfect tinder, spinifex gets a 9), one match will often
start a raging bonfire. Spinifex has been called an ideal pyrophyte.
Spinifex is an exceedingly important habitat element for many species of
Australian desert lizards. Individuals of most species take refuge in spinifex
from predators and the sun. Some species occur only in large long unburned tussocks.
 
Martu hunter burning spinifex to find Sand Monitor lizards Varanus gouldii (Photo courtesy Rebecca Bliege Bird) Most fires are started by lightning, and can burn for weeks across thousands of square kilometers of uninhabited desert. Australia is probably the last place on Earth where wildfires are allowed to rage completely out of control. Wildlife managers in Australia try to optimize fires to create a fine-grained patchwork of habitats of different ages. Fires vary considerably in intensity as well as extent. In 1992-94, we undertook a detailed landscape-level remote sensing study of wildfires as agents of disturbance promoting habitat diversity in the Great Victoria Desert of Western Australia using satellite imagery (Haydon et al. 2000a, 2000b). About 450 fires occurred in each of two Landsat scenes over the 20 year period from 1972 to 1992, but we found very little reburn. Fire return interval for vegetation was estimated to be about 20 years. Most fire studies use a chronosequence method using areas known to have burned at different times (e.g., see Masters 1996. This allows many years worth of data to be collected during a short time. While useful, this method has clear limitations because of confounding factors involving differences among study plots. Differences between recently burned and long since burned plots may have different biological properties for many reasons; including soil chemistry, topology, or even proximity to other sites with very different physical or biological properties. To avoid such problems, I undertook a longitudinal long-term study at one site, although, of course, limits on manpower dictated that it could not be replicated.  Following a fire, spinifex rejuvenates rapidly from live roots as well as by seedling
establishment. Newly-burned areas are very open and consist mostly of bare ground
covered with tiny, well-spaced, clumps of spinifex. Unburned patches, in contrast,
are composed of large ancient tussocks, frequently close together with little or no
open space between them.
Effects on lizards and lizard microhabitats are drastic, yet vary in time and location.
Many or even most individual lizards live through the burn itself, only to succumb in
the fire-reduced habitats and microhabitats that can last for years. Some lizard species
with open habitat requirements reinvade and repopulate burned areas rapidly. Other
species that require large spinifex tussocks for microhabitats often become very rare
or even vanish altogether after a burn. However, such 'climax' species (those living
in long-undisturbed habitat) can continue to exist in isolated pockets and patches of
unburned habitat. Fires thus create a habitat patchwork, increasingly recognized as a
major contributor to species richness (Pianka 1989, 1996). Indeed,
wildlife managers in Australia use controlled burns to attempt to maintain an appropriate
fine-grained mixture of habitats of different ages since fire.
Following a fire, spinifex rejuvenates rapidly from live roots as well as by seedling
establishment. Newly-burned areas are very open and consist mostly of bare ground
covered with tiny, well-spaced, clumps of spinifex. Unburned patches, in contrast,
are composed of large ancient tussocks, frequently close together with little or no
open space between them.
Effects on lizards and lizard microhabitats are drastic, yet vary in time and location.
Many or even most individual lizards live through the burn itself, only to succumb in
the fire-reduced habitats and microhabitats that can last for years. Some lizard species
with open habitat requirements reinvade and repopulate burned areas rapidly. Other
species that require large spinifex tussocks for microhabitats often become very rare
or even vanish altogether after a burn. However, such 'climax' species (those living
in long-undisturbed habitat) can continue to exist in isolated pockets and patches of
unburned habitat. Fires thus create a habitat patchwork, increasingly recognized as a
major contributor to species richness (Pianka 1989, 1996). Indeed,
wildlife managers in Australia use controlled burns to attempt to maintain an appropriate
fine-grained mixture of habitats of different ages since fire.
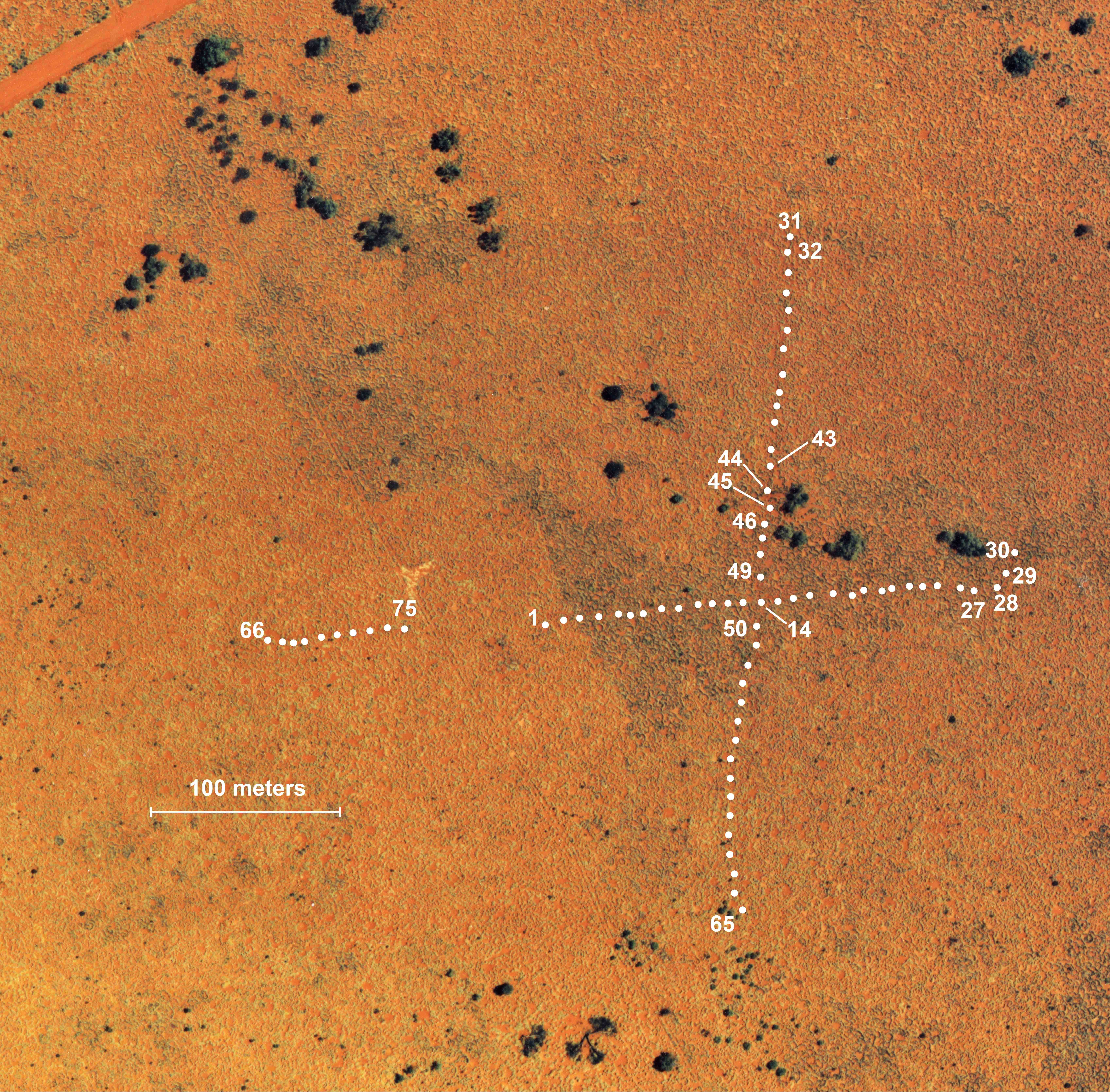
In the Austral Spring of 1992, I decided to undertake an imperfect experiment, without replication, and set up a new study site, the B-area, with pit trap lines and drift fences, monitoring it before and after a controlled burn, shown in the above high resolution aerial photograph taken on 17 February 1993. Positions of 75 pit traps are shown with solid white circles, along with a scale. Hundreds of termitaria, circular open areas about the same size as pit trap circles, are scattered about the image. Notice the patches of older long unburned spinifex embedded within larger patches burned more recently. The main east-west track from Laverton WA to Coober Pedy SA via Neale Junction is in the northwest (upper left hand) corner. Pre-burn (long unburned) pit trap samples were collected for 3 months in 1992. A controlled burn was undertaken at this study site in October 1995 and the site has been monitored at regular intervals following the fire during 1995-96, 1998, 2003, and 2008 to assess changes in community structure. Lizard censuses from an unburned site 4 km north of the experimental burn site (the R-area) can be considered a partial control, as they provide a reference for understanding population and community change in relation to fire on the B-area. 

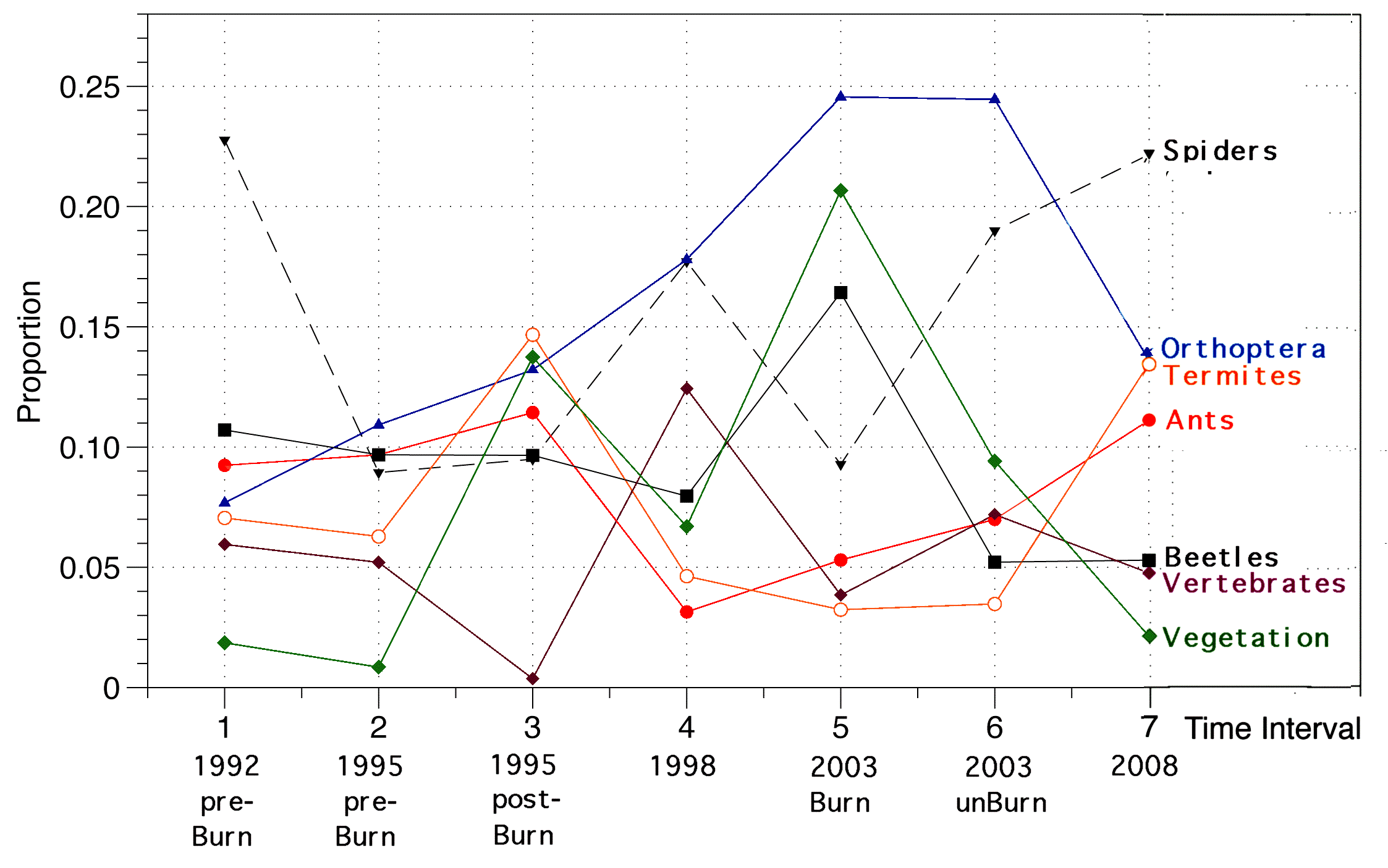
A few dozen scattered tussocks escaped the fire, providing limited refuges for surviving lizards. Immediately after the controlled burn, the area was walked searching for lizard fire victims but very few were found. Most lizards survived the fire by escaping into burrows. Pits were opened without drift fences for one month in late 1995 and again for two weeks in January 1996. The seven most abundant lizard species were two agamids Ctenophorus isolepis and Ctenophorus nuchalis, and five Ctenotus skinks, C. ariadnae, C. calurus, C. hanloni, C. pantherinus, and C. piankai. Abundances varied considerably through the course of the fire succession cycle. Most striking, abundances of the two agamids C. isolepis and C. nuchalis were negatively correlated, fluctuating out of phase inversely with each other, with C. nuchalis reaching very high population densities immediately after each burn, whereas C. isolepis densities diminished after fires and were more abundant in unburned or recovering vegetation cover. Lizard faunas and foods eaten on the B-area have been examined at 7 different time intervals in the fire succession cycle from original long-unburned to 16 years post-burn to better understand the dynamics of this ecosystem. These long-term census data provide valuable insights into how individual species respond (species composition, relative abundance, dietary flexibility, and reproductive tactics). A total of 2872 individual lizards representing some 46 species have been collected at the B-area from 1992 through 2008.
|
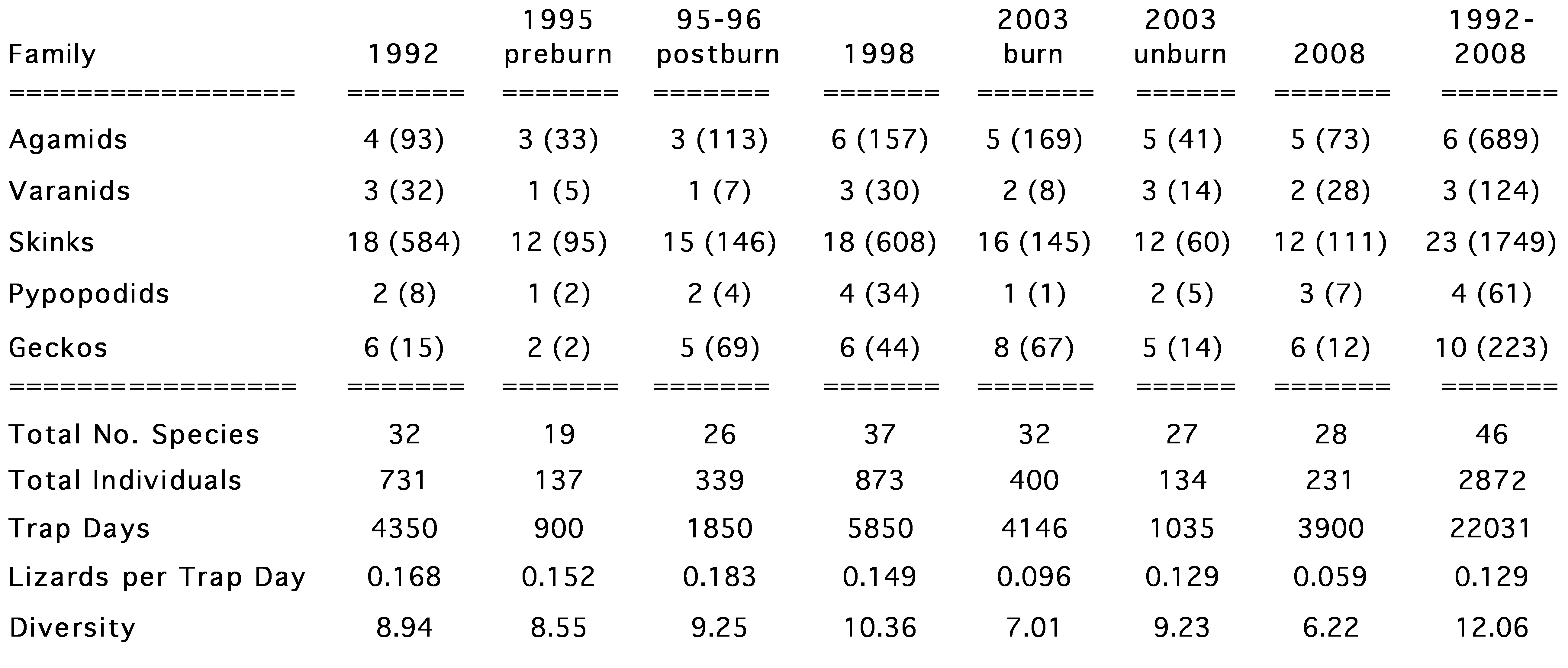
|
Diversity on the B-area peaked in 1998, a few years following the 1995 controlled burn, but has fallen since then. Pit trapping results from the same pits sampled at the same time of year (Sept.-November, Austral Spring) from 1998 to 2003 to 2008 demonstrate decreases in numbers of individual lizards as measured by trap success (numbers of lizards captured per trap day). Numbers of species and species diversity also decreased. Some species appear to have diminished in abundance, while others may have gone extinct during this decade. Although thousands of individual lizards were removed, new animals moved in from vast adjacent areas as fast as residents were trapped -- plots of trap success versus time during a given sampling period fluctuated with weather, but did not trend downwards. The 3-5 year intervals between sampling periods are ample for populations to reproduce and recolonize. The netted dragon C. nuchalis was rare initially during pre-burn censuses in 1992 and in September-early October 1995, but increased massively in abundance immediately after the burn in October 1995. This increase in numbers was too rapid to have been a result of reproduction, but rather appears to have been due to immigration into the area (an alternative, untestable, hypothesis is that these lizards were present before the fire but did not fall into pit traps until after the burn opened up the vegetation). An unexpected natural wildfire burned part of the study site in 2000, clearing the vegetation, which allowed C. nuchalis to increase again in 2003. With heavy rainfall, spinifex again recovered rapidly over the next 5 years, greatly reducing C. nuchalis numbers again in 2008. C. isolepis became more abundant as C. nuchalis numbers decreased. Relative abundances of the 5 species of Ctenotus skinks behaved similarly to those of the agamid Ctenophorus isolepis, with which species their abundances are positively correlated. Three species of nocturnal geckos, Diplodactylus conspillatus, D. damaeus, and Rhynchoedura ornata, also increased in abundance following fire. 
The 2000 natural fire burned only a portion of the B-area, leaving some traps in the unburned maturing vegetation and others in the recently burned, open habitat. This unexpected occurrence was due to high rainfall, rapid regeneration of spinifex from roots, and suggests that fire return interval has shortened. It also allowed for continued tracking of part of the recovering habitat in addition to replicating a sample of the original imperfect experiment. Three species increased in abundance immediately after each fire, the geckos Rhynchoedura ornata and Diplodactylus damaeus, and an agamid Ctenophorus nuchalis -- all three decreased rapidly after fires as spinifex recovered and open space diminished. 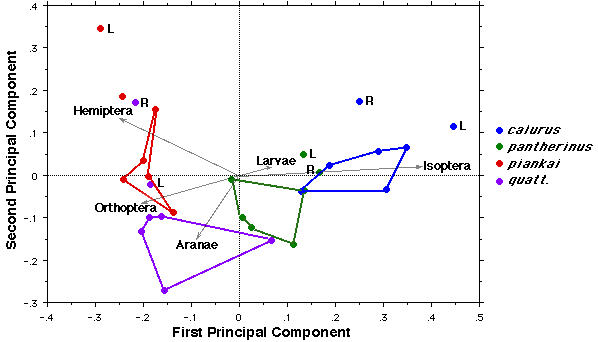
 The following figure plots the first two principal components of dietary niche space, showing overall diets of all species over the course of the 16-year fire succession cycle. These two principal components capture 60% of total variation in diet. Positions of food categories are shown in red with the length of vectors proportional to relative importance. Dark arrows show that diets move counterclockwise through these 2 dimensions, from 1992 to 1995 pre-burn to 95-96 post-burn to 2003 re-burn to 2003 unburned to 1998 and then to 2008 when diets returned to near 1992 pre-burn conditions.  References Goodyear, S. E. and E. R. Pianka. 2011. Spatial and temporal variation in diets of sympatric lizards (genus Ctenotus) in the Great Victoria Desert, Western Australia. J. Herpetology 45: 265-271. Read On Line. Download pdf Haydon, D. T., J. K. Friar, and E. R. Pianka. 2000a. Fire Driven Dynamic Mosaics in the Great Victoria Desert I: Fire Geometry. Landscape Ecology 15: 373-381. Download pdf. Haydon, D. T., J. K. Friar, and E. R. Pianka. 2000b. Fire Driven Dynamic Mosaics in the Great Victoria Desert II: A spatial and temporal landscape model. Landscape Ecology 15: 407-423. Download pdf. Masters, P. 1996. The effects of fire-driven succession on reptiles in spinifex grasslands at Uluru National Park, Northern Territory. Wildlife Research. 23: 39-48. Pianka, E. R. 1989. Desert lizard diversity: additional comments and some data. American Naturalist 134: 344-364. Download pdf. Pianka, E. R. 1996. Long-term changes in Lizard Assemblages in the Great Victoria Desert: Dynamic Habitat Mosaics in Response to Wildfires. Chapter 8 (pp. 191-215) in M. L. Cody and J. A. Smallwood (eds.) Long-term studies of vertebrate communities. Academic Press. Download pdf. Pianka, E. R. and S. E. Goodyear. 2012. Lizard responses to wildfire in arid interior Australia: Long-term experimental data and commonalities with other studies. Austral Ecology 37: 1-11. Download pdf |