|
Landscape ecology and ecological biogeography (For an outline, click here) 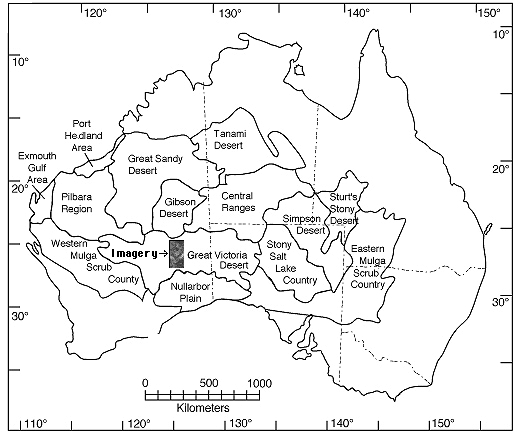
Fire Succession in Inland Western Australia The importance of spatial scale has been neglected in traditional ecology, although not in the emerging field of landscape ecology. While the implications of the landscape on ecology have long been appreciated, only recently have quantitative methods of study been exploited. In the past, ecologists, including myself, have focused on local-level processes. Larger scale regional factors also control local phenomena. Local species richness may often be a consequence of regional processes. Relatively little empirical attention has been given to the interaction between these two levels. Unfortunately, few complete closed regions remain unfragmented by human activities in which regional and local phenomena can still be studied simultaneously. I am undertaking such a study in the uninhabited Great Victoria desert of Western Australia, an area with an extremely high diversity of lizards. Fires were once a major agent of disturbance in all grassland and semidesert biomes, including the North American tall grass prairies. Most of these ecosystems have now been reduced to mere vestiges, and controlled burning and/or fire control are practiced by humans almost everywhere. The inland Australian desert is one of the last remaining areas where wildfires remain a regular and dominant feature of an extensive natural area largely undisturbed by humans. An important fire succession cycle, which generates spatial and temporal heterogeneity in microhabitats and habitats, is evident in this region. These regional processes promote local diversity. This system is being studied at the local level in the field in Australia and at the regional level at the University of Texas using aerial photos and multispectral satellite imagery. High resolution satellite imagery of these areas, which has been collected since 1972, offers a powerful way, heretofore underutilized, to acquire regional level data on the frequency and phenomenology of wild fires, and thus the system-wide spatial-temporal pattern of disturbance. Much of the digital satellite data has been acquired by Landsat, archived on magnetic tape. A complete analysis of wild fires for the Great Victoria desert region requires about one hundred images. Imagery has been purchased and analyzed to detect burned areas. Spectral and spatial statistics have been computed for nearly a thousand fires, and the probability that a given area will burn has been estimated. Other data collated for each fire include: date, location, area, perimeter, compass direction (of burn and prevailing wind), ground cover characteristics, extent of reticulation, as well as various fractal dimensions. (To read more about the remote sensing study, click here) Age and size distributions of burn patches have been estimated. Eventually, I hope to acquire supporting imagery from other grassland areas, particularly the Kalahari semidesert of southern Africa, will be acquired and used for comparative purposes (fires in the Kalahari do not appear to reticulate to as great an extent as they do in Western Australia).
Field work is currently underway on the ground to document rates of closure of spinifex, to map vegetation structure, and to determine which animal species are present, and in what abundance, at various stages following burns. Low level aerial photography has been acquired and will be digitized, georeferenced, and analyzed to make detailed maps of vegetation structure for use in computer simulations of fire dynamics. Vertebrate faunas and insects, particularly foods eaten by lizards, will be compared at recently burned sites with those at various stages of post fire recovery to collect preliminary data for modelling aspects of the dynamics of the fire succession cycle. A mature spinifex site was monitored for several years prior to burning and will be censused over the long-term as often as possible post-burn. Fires are currently being simulated on the computer to mimic observed fire geometry. Precipitation, rates of accumulation of combustibles, and the insect and vertebrate faunas of these study sites will be monitored as often as possible over the next two decades to collect more precise data on the fire succession cycle. A major goal of this study is to obtain baseline data on temporal patterns, spatial structure, and distribution of disturbances. These data will form the backdrop for a more detailed study of the population dynamics and dispersal abililties of component species. Such data on the component species will be fitted into the overall spatial-temporal mosaic in an effort to explain the persistence of this diverse desert fauna. Ultimately, I plan to model the entire Great Victoria desert region as a dynamic habitat mosaic so as to understand mechanisms of coexistence of its component species and the effects of fire disturbance in maintaining lizard diversity in this region. 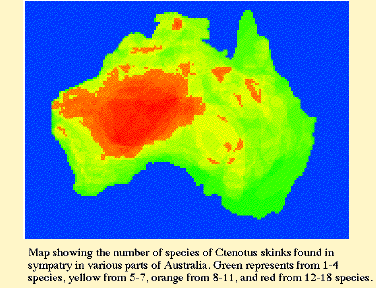 Most genera contain relatively few species. The Australian skincid genus Ctenotus is an exception having undergone a very extensive adaptive radiation within the last 4-10 million years. It is the most speciose lizard genus (about100 species) in Australia and among the top five species rich lizard genera in the entire world (others are Anolis, Lerista, Liolaemus, and Sceloporus). The phylogeny of Ctenotus will be recovered by sequencing DNA from mitochondrial and nuclear genes that are evolving at varying rates. Frozen tissues of 75 species are already on hand. A phylogeny of about 60 species of Ctenotus will be recovered which will allow us to test the monophyly of the genus and current phenetic groupings within Ctenotus. Species groups will be redefined and, in all probability, new species will be discovered.  Once a phylogeny is in place, we will examine extensive ecological information on 14 species in the Great Victoria desert region within a phylogenetic perspective to test various hypotheses about evolution and biogeography in this diverse genus. Traits that have evolved rapidly among species within this genus include toe lamellae, body temperatures, habitat specificity, dietary and microhabitat niche breadth, head length, relative hindleg length, body size, variability in selected morphological traits, extent of geographic range, clutch size and other reproductive parameters such as relative clutch mass and size of oviductal eggs (expenditure per progeny). Anatomical and ecological convergences will be elucidated using phylogenetic systematics, modern comparative methods, and phylogenetically-based, multivariate statistical procedures. The extent to which phylogeny has constrained morphology and ecology will be ascertained. Historical biogeographic analysis will shed light on the origin and radiation of various species groups within Ctenotus, the aridification and vicariance of Australia, and will bear on the origin of the Australian arid zone fauna as a whole. Such a comprehensive study of a speciose genus will elucidate the factors that allowed this major adaptive radiation. It will also enable prediction of the probable ecologies of unstudied species of Ctenotus, which will then be tested by direct examination in the field.  Ecological Biogeography Latitudinal gradients in species diversity Comparisons of species densities of North American and Australian vertebrates Correlations with the physical environment (precipitation, sunshine) Complementarity Nocturnal insectivores: frogs vs. lizards, Old World vs. New World (To read about ecomorphology, click here) References on Ecological Biogeography Colwell, R.K. and D.W. Winkler. 1984. A null model for null models in biogeography. Chapter 20 (pp. 344-359) in D.R. Strong, D. Simberloff, L.G. Abele and A.B. Thistle (eds.), Ecological communities: Conceptual issues and the evidence. Princeton University Press, Princeton, N.J. Duellman, W. E. and E. R. Pianka. 1990. Biogeography of Nocturnal insectivores: Historical eventts and ecological filters. Ann. Rev. Ecol. Syst. 21: 57-68. Keast, A. (ed.) 1981. Ecological Biogeography in Australia. D. W. Junk, The Hague, Netherlands. MacArthur, R. H. 1972. Geographical Ecology. Harper and Row, N.Y. 269 pp. MacArthur, R. H., and E. O. Wilson. 1963. An equilibrium theory of insular zoogeography. Evolution 17: 373-387. MacArthur, R. H. and E. O. Wilson. 1967. The Theory of Island Biogeography. Princeton University Press, Princeton. Pianka, E. R. 1986. Ecology and natural history of desert lizards. Analyses of the ecological niche and community structure. Princeton University Press, Princeton, N.J. Pianka, E. R. and J. J. Schall. 1981. Species densities of terrestrial vertebrates in Australia. Chapter 59 (pp. 1675-1694) in Ecological Biogeography in Australia, A. Keast, ed. D. W. Junk, The Hague, Netherlands. Schall, J. J. and E. R. Pianka. 1978. Geographical trends in numbers of species. Science 201: 679-686. Winemiller, K. O. 1991. Ecomorphological diversification in lowland freshwater fish assemblages from five biotic regions. Ecological Monographs 61: 343-365. Back to Herpetology Home Page Pianka lab page |

